reproductives.htm
by George H. M. Lawrence, Professor of Botany at the Bailey Hortorium, Cornell University, 1951
Photo-copied by Maung Kan Tun from the original text.
Edited and set in HTML by U Kyaw Tun (M.S., I.P.S.T., USA),
Tun Institute of Learning (TIL) and the staff of TIL.
Not for sale. No copyright. Free for everyone.
Prepared for students and staff of TIL
Computing and Language Center, Yangon, MYANMAR :
http://www.tuninst.net ,
http://www.softguide.net.mm ,
www.romabama.blogspot.com
index.htm |
Top
Lawrence-indx.htm
Principles of Taxonomy
Reproductive elements
Reproductive elements
and their taxonomic significance p064
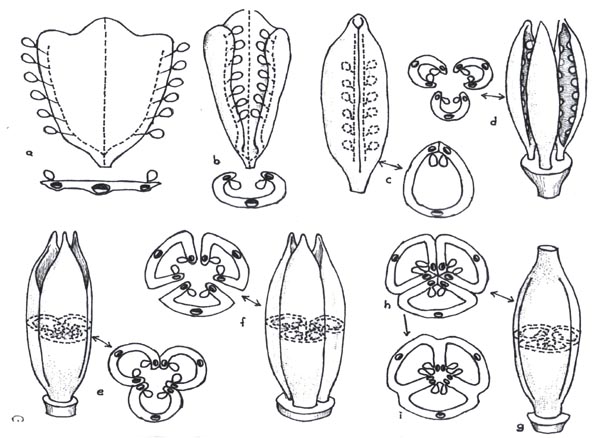
Flower
Perianth -- the envelope: sepals; petals
p066
Male reproductive
organs: stamen p067
Fig. 4. Androecium and stamen types p070
Female
reproductive organs: carpel p072
Fig. 5. Gynoecial types p073
Fig. 6 Hypothetical evolution of an ovary
p074
Placentation-types
p075
Fig. 7. Presumed evolution of placentation
types p076
Ovary position p079
Fig.8. Ovary position p080
Fig.8. Ovary position p080
04.4 Flower types p081
Fig. 9. Flower types and arrangement p082
Distribution of
sexes p.082
Main Index of DB |
Top
Index of Collected papers
Index of Lawrence's Taxonomy
Contents of this page
[{cont. p064}]
Taxonomists, since and before the time of Linnaeus, have relied heavily on the value of characters inherent in the reproductive parts of vascular plants to serve as criteria of particular significance in their classifications. In the pteridophytes reproductive characters are associated in general with the sporangia and sex organs of the gametophyte; in the gymnosperms they are those of the pollen-producing and fruiting structures; and in the angiosperms are those of both the flowers and the fruiting structures. This last taxon is the most complex of all, and phylogentically is the most highly developed and advanced. For these reasons great diversity and complexity of reproductive characters are typical of its members. Since the objectives of taxonomy include identification and classification, it is necessary to understand the composition of the flower, of the theories purporting to account for the origin of its components, and of the presumed evolutionary levels represented by present-day types of these components. The disciplines of morphology, anatomy, and paleobotany have contributed the data and the conclusions responsible for these understandings.
Prerequisite to any basic understanding of the flower is an appreciation of the view that vascular plants structurally are evolved from or composed of different types of branch systems. As applied to flowering plants,
Lawrence 065
and on the evidence adduced from the fossil record that terminal reproductive structures were present prior to the evolution of foliaceous leaves, it is probable that at least 3 types of branch systems have served as antecedents of present-day leaflike structures (Wilson, 1941).
These 3 types are:
1. a branch or cluster of branches bearing terminal sporangia from which the
stamen is descended;
2. branches which became flattened (but were never
primarily photosynthetic) to become the ovule-bearing
carpels;
3. sterile
branches or branch systems which became the leaves.
This postulate should be kept in mind when considering the conventional and very generally accepted theory (Eames, 1931) that "the flower morphologically is a determinate stem with appendages, and these appendages are homologous with leaves." In other words, flowers are homologous with leafy stems; the pedicel and receptacle of the flower are homologs of the stem axis, and the sepals, stamens, and pistil (s) are homologs of the leaves (i.e., the latter are appendages of the stem). If the quotation form Wilson is accepted, it becomes clear that although foliaceous leaves may be appendages of stems they may be presumed to have been evolved by the dorsiventral compression and lateral fusion of photosynthetic branches.19 (fn065-19) Similar origins were postulated by Wilson to account for the leaflike appendages represented by the stamens and by the carpels. Recognition that the foliar units are fundamentally modified branch systems brings the contentions of some botanists that floral parts are of appendicular origin into complete harmony with the contention of others that they are of stem or branch origin. Statements to the effect that foliage leaves and floral elements are homologs does not mean that they are necessarily derived one from the other.20 (fn065-20)
UKT: The flower, whether immature (bud) or mature is made up of 2 parts: the "envelope" and "contents" (part or parts to be protected by the envelope). The envelope is the perianth, and the contents the generative organs (androecium - male organs, and gynoecium - female organs)
[Go back flo-call1-b; stamen-call1-b]
The flower typically is composed of the following 4 types of components (as
arranged from the bottom of the floral axis upwards):
1. sepals, collectively the
calyx;
2. petals, collectively the corolla;
3. stamens, collectively
the androecium; and
4. carpels (the basic unit of the pistil), collectively
Lawrence 066
the gynoecium. The calyx and corolla comprise the perianth (floral envelope, perigone); neither is essential to the functioning of the flower as a reproductive structure.
UKT: The description of the flower given above is that of a mature flower. In a juvenile flower or bud, there are only 3 parts: the outer covering (proto-calyx), the inner covering (proto-corolla), and the immature sex (both male and female) organs.
See androecium and gynoecium.
UKT: see peri-
The perianth is a non-essential as a reproductive structure.
The perianth is an accessory part of the flower more or less enveloping the organs of reproduction, and is classically treated as composed of an outer cycle or whorl (the calyx) and an inner cycle or whorl (the corolla). This explanation is one of extreme simplicity. The origin of the perianth of the modern flower is usually explained by one of two hypotheses.
By one interpretation, the flower is assumed to have primitively been provided
with a perianth of 2 cycles (one the calyx and the other the corolla). If it is
assumed that upon phylogenetic reduction the corolla inevitably drops out first
and the flower then becomes apetalous, the remaining perianth is one of sepals.
On the other hand, if it is accepted that reduction may affect each cycle
equally, and that the corolla is capable of assuming a sepaloid form (or the
sepals a petaloid form), then a perianth is to be considered composed of units
of undeterminable phyletic orgin with its components termed tepals (as in
many monocots).
The second and more widely accepted interpretation treats the ancient flower to
have been an axis bearing many foliaceous bracts in a spiral arrangement, and
assumes that the apical ones were fertile (mega- and microsporophylls) and the
lower ones were sterile. These sterile bracts were similar in appearance, with
the upper ones gradiently resembling more closely the microsporophylls than
they did the lowermost sterile bracts. Because of this foliaceous similarity
they are designated tepals. From this primitive ancient condition, and in
conformance with the laws of phyllotaxy interacting with progressive spatial
restrictions (resulting in apparent vertical compression of the axis), there was
evolved the modern flower. This has a perianth that is usually dicyclic (and
sometimes tricyclic) and pentamerous,
tetramerous, trimerous, or dimerous.
By this view, all parts of the perianth are homologous, and phylogenetically are
tepals. The terms calyx and corolla become descriptive terms of taxonomic
importance, but may lack phylogenetic significance. Differentiation into calyx
and corolla may and probably did take place at many phylogenetic levels in
angiosperm evolution, and there is ample evidence of petaloid calyces and of
greater significance to plant identification than to classification. Some
apparent corollas have been derived from the andeoecium (Aizoaceae), other
represent petaloid calyces (many ranaliaceous plants).
Lawrence 067
Sepals. The sepals compose the outermost series of the components of the typical flower and collectively comprise the calyx. Most sepals are supplied by three vascular strands (traces or bundles), as are also most leaves, and possess anatomy like that of the leaves of the plant on which they appear. For these reasons, they have been considered morphologically to be bracts and to have been derived from foliaceous leaves. Generally they are green in color (photosynthetic) and bear a resemblance to bracts or leaves. They may be distinct21 (fn067-21) (polysepalous or aposepalous) or connate (gamosepalous or synsepalous) or in some families be reduced to modified hairs or seales and referred to collectively as a pappus.22(fn067-22) Sometimes they are highly colored and petallike (Delphinium, Helleborus, Mirabilis)
Petals. The petals compose the second series, or inner envelope, of the perianth (i.e., the corolla). Morphologically and anatomically they are more often like sterile stamens than they are like leaves. Evidence for this view is based on the presence (in most petals) of a single vascular strand, a characteristic also of most stamens. Usually the petal is colored other than green (nonphotosynthetic) and contributes to the showiness of the flower. It has been considered that corollas of distinct petals are more primitive than those whose petals are marginally connate, and this situation has been accorded much significance in classification schemas. However, Hutchinson (1948), who accepted polypetaly (apopetaly) as more primitive than gamopetaly (sympetaly), rejected the view that these two criteria were of sufficient phyletic importance to separate orders of plants into two subclasses (Polypetalae and Gamopetalae). The criteria were treated by him to be of minor phyletic importance. Terms used in describing the shape, texture, and disposition of perianth parts are the same as are used in describing similar parts of leaves.
Stamens.
The stamen is the male reproductive organ of the flower.
Collectively the stamens comprise the
androecium. The stamen is conventionally
treated as composed of an anther (the pollen-producing element) and a filament
(the stalk). The more recent views of Wilson (1941), based on anatomical studies
of the androecium of the Parietales and
Lawrence 068
Malvales, differ in some respects from the perhaps better-known classical theories of androecial origins as presented originally by Goethe and by de Candolle (1813). The older theories treated the stamen as a sporophyll, a fertile homolog of the foliaceous leaf, whereas by Wilson's view it was treated as a direct derivative of a primitive branch system bearing terminal sporangia, that is, a branch system that did not pass through any stage of dorsiventral compression is its phyletic development. In amplifying this premise, Wilson further considered that :
A hypothetical primitive stamen may be conceived, in which a relatively long shank, or axis, itself an arm or limb of a dichotomy, is terminated by a system of dichotomously divided branches, each ultimate branchlet bearing a single sporangium. By reduction of the two most remote dichotomizing branches, with the resultant fusion of pairs of sporangia, an anther of two synangia each composed of two sporangia may be derived. . . The evolution of anthers, two-celled at maturity, but four-celled in ontogeny, probably took place very early in the evolution of the Angiosperms.
. . . it may be concluded that in those flowers with numerous stamens, . . . the stamens represent not primary organs or sporophylls of the flower, but the apices of branch systems, the bases, or many branches of which, have dropped out in the course of evolution. The presence of a few and fixed number of stamens is then to be regarded as a modification of the condition obtaining in the flower with numerous stamens, each stamen representing the reduction of an entire branch system.
This view of the origin of the stamen is pertinent also to any consideration of the origin of the petal of most petialiferous families, since the petals are generally conceded to have been evolved from stamens. The stamens and petals of most flowers are supplied by a single trace, but in members of several families (including the Magnoliaceae and Lauraceae) each stamen is supplied with 3 traces evidence of a primitive staminal condition in these representatives. It was postulated by Wilson that the staminal tube present in monadelphy (mondelphous stamens being those whose filaments are fused laterally into a cylinder, as in the hollyhock) is the "result of fusion of several branch systems" and is indicative of a primitive androecial condition.
If the above hypotheses are accepted, monadelphous stamens are among the more primitive, diadelphous stamens slightly more advanced, while those groups having stamens clustered in fascicles are more primitive than distinct stamens. Among the groups with androecia of distinct stamens, those with stamens numerous would be considered more primitive than those with stamens few and definite in number, and androecia reduced to a single stamen would be the most advanced of all types.
It is significant that angiosperm families now generally conceded to be
Lawrence 069
the more primitive (Magnoliaceae, Ranunculaceae) have numerous stamens that are spirally arranged. Furthermore, since the stamens are distinct and free, it is to be presumed (in accordance with Wilson's findings) that they represent emerging branch tips of complicated branch systems. The spiral arrangement of floral parts has been accepted as a condition more primitive than that of whorled arrangement. A study of androecial types indicates that the views of Wilson are not necessarily contradictory to the classical concept that spirally arranged stamens are more primitive than whorled stamens. Wilson has attempted to account for situations where there exists a reduction in the number of stamens (per unit branch of the androecium). This reduction would appear to be an independent line of advance from that represented by the reduction from the spiral to the whorled arrangement. There is anatomical evidence that :
1. some present-day stamens, superficially simple, are individually the remnants of much reduces (and usually dichotomous) branch systems (e.g., in the Melastomaceae),
2. within some orders of plants monadelphy preceded polyandry (e.g., Malvales),and
3. that in some admittedly primitive orders the androecium of present-day plants is represented by tips of emerged branch tips disposed in a high phyllotaxy (i.e., spirally arranged, as the stamens in Magnoliales and Ranales).
The fossil record produces no known barrier to the view that each of these 3 manifestations may have arisen independently. Each of the 3 may represent lines of androecial evolution whose phyletic significance can be evaluated only in relation to order phyletic criteria. A taxon accepted to be advanced, as determined by an aggregate of characters, may possess an androecium of a presumed primitive type (as in Malvaceae).
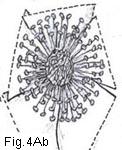
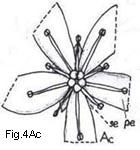
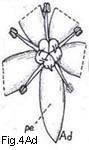
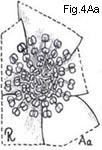 Relatively few families are characterized by having spiral stamen arrangements
(Fig. 4 Aa), and in the majority of families the stamens are whorled (Figs. 4 Ab-Ad).
Families with a generally fixed number of whorled stamens usually can
Relatively few families are characterized by having spiral stamen arrangements
(Fig. 4 Aa), and in the majority of families the stamens are whorled (Figs. 4 Ab-Ad).
Families with a generally fixed number of whorled stamens usually can
 be divided into those whose stamens are in two whorls or are in a single whorl. Presence of
a single whorl is considered the more advanced condition because, in most
instances, the anatomical evidence has shown that the second (and inner) whorl
of stamens that were present in their ancestral types has since been lost by
suppression or by reduction. In some taxa, the remnants of this second whorl can
be recognized as staminodes or nectaries, or as additional petals. The order or
sequence in which stamens develop is of taxonomic importance, especially in an
androecium of many stamens. In most families development is
centripetal (the
outer ones developing before the inner, sometimes described as
acropetal) but in
others it is centrifugal
be divided into those whose stamens are in two whorls or are in a single whorl. Presence of
a single whorl is considered the more advanced condition because, in most
instances, the anatomical evidence has shown that the second (and inner) whorl
of stamens that were present in their ancestral types has since been lost by
suppression or by reduction. In some taxa, the remnants of this second whorl can
be recognized as staminodes or nectaries, or as additional petals. The order or
sequence in which stamens develop is of taxonomic importance, especially in an
androecium of many stamens. In most families development is
centripetal (the
outer ones developing before the inner, sometimes described as
acropetal) but in
others it is centrifugal
Lawrence 070
(basipetal).23(fn070-23) This distinction indicates relationships between groups of one type or the other, and the opinion is held (Corner, 1946) that "the primitive massive, centrifugal androecium must have been derived from the usual centripetal state."
In addition to the presumed phyletic significance and origin of stamens as discussed above, there are numerous other characteristics associated with them that are of significance especially in the taxonomy of minor
Fig. 4. Androecium and stamen types
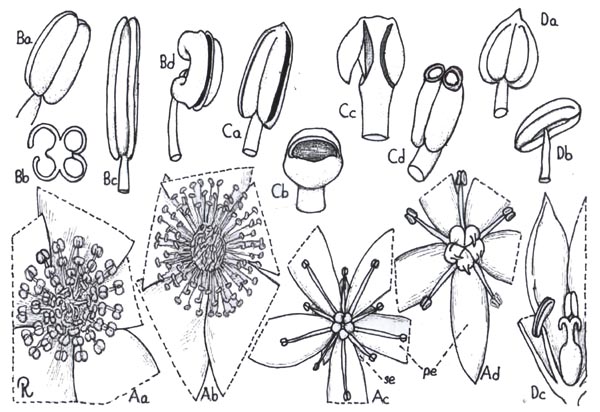
Fig. 4. Androecium and stamen types. A, androecium types: Aa, stamens spirally arranged; Ab, stamens in three whorls; Ac, stamens in two whorls, those of outer whorl the shorter (diplostemonous); Ad, stamens in a single whorl (dehiscence introrse). B, anther cell number: Ba, cells four at anthesis; Bb, same cross-section: Bc. cells two at anthesis; Bd, cell one at anthesis. C, anther dehiscence types; Ca, longitudinal; Cb, transverse; Cc, valvular flaps; Cd, apical pores, D, anther position; Da, basifixed; Db, dorsifixed and versatile; Dc, dorsifixed, not versatile (dehiscence extrorse).
taxa. The anther of the typical stamen at maturity (anthesis) is composed of two thecae (actually each theca at time of anthesis represents two sporangia, the theca often termed a cell or chamber) separated from one another by an extension of the filament passing between them (connective). Fundamentally the anther is of 4 thecae, but the septum between the two adjoining ones situated on the same side of the connective usually disappears during the late ontogenetic development of the anther and prior to anthesis, although in a few taxa (as Lauraceae) the septa persist and the anthers are then 4-thecate at anthesis. In some families (Malvaceae, some Bombacaceae) the anther has a single theca, a situation
Lawrence 071
shown by Wilson to be due to "the fusion of two neighboring sporangia into
single synangia." The anther is fundamentally an organ that is terminal on the
filament and when so it is said to be basifixed. A more advanced
situation is present when the anther appears to be attached
 dorsally to the
filament and is then said to be dorsifixed, and when dorsifixed it is
sometimes versatile (Fig. 4 D). The anther cell (theca) dehisces
typically by a vertical or longitudinal split, but in more advanced situations
the dehiscence may be transverse (Hibiscus, Elatine) or by a pore
(Polygalaceae, most Ericaceae). In some families the connective between the
thecae becomes highly developed and appendaged, even to the extent (in Cannaceae
and Marantaceae) of being petaloid and showy. The androecium is said to be
syngenesious when its anthers are connate marginally. The anthers of syngenesious androecia ordinarily ( in Campanulaceae spp., Compositae) form a
cylinder through which the style or stigmas pass, but sometimes (in some
Gesneriaceae) the syngenesious anthers are dissociated and remote from the style
(see Fig. 317e).
dorsally to the
filament and is then said to be dorsifixed, and when dorsifixed it is
sometimes versatile (Fig. 4 D). The anther cell (theca) dehisces
typically by a vertical or longitudinal split, but in more advanced situations
the dehiscence may be transverse (Hibiscus, Elatine) or by a pore
(Polygalaceae, most Ericaceae). In some families the connective between the
thecae becomes highly developed and appendaged, even to the extent (in Cannaceae
and Marantaceae) of being petaloid and showy. The androecium is said to be
syngenesious when its anthers are connate marginally. The anthers of syngenesious androecia ordinarily ( in Campanulaceae spp., Compositae) form a
cylinder through which the style or stigmas pass, but sometimes (in some
Gesneriaceae) the syngenesious anthers are dissociated and remote from the style
(see Fig. 317e).
In the development of the anther there are produced in the pollen sac many usually rounded cells termed pollen mother cells. Four pollen grains usually are formed from each of these. Pollen grains are of different morphological types and are receiving increasing recognition as a source of valuable taxonomic characters. The outer membranous coat of the pollen grain is cuticular in composition and is termed the exine. Often it is formed into various kinds of sculptured surfaces (furrowed, papillose, honeycombed, echinate, etc.) that are characteristic of particular taxa. Pollen grains that are free from one another are said to be granular. Sometimes the pollen grains are in clusters of 4, tetrads (as in many Ericaceae), but in at least two families (most Asclepiadaceae, Orchidaceae) the pollen of each anther sac is agglutinated into usually waxy to firm masses called pollinia. Pollen grains are often of value in distinguishing possible or suspected hybrids from nonhybrids, since the pollen of some hybrids is nonviable and is shrunken and nonturgid in appearance.24(fn071-24)
A conspicuous modification of the androecium is the reduction of one or more stamens to sterile structures termed staminodes. These may
Lawrence 072
appear as a filament minus an anther (Penstemon), as expanded petal-like structures (members of the Aizoaceae, in Canna), knobbed glandular processes (many orchids), or in the form of a nectary (Loasaceae).
Some groups of plants are pollinated almost exclusively by wind and are described as being anemophilous. Others are pollinated largely by animal vectors, particularly by insects, and are said to be entomophilous. There has been considerable discussion over which of these conditions is the more primitive, and some of the basis for divergent views may be due to the probability that some of the present-day anemophilous plants perhaps have evolved from ancestors that were entomophilous, and that the latter in turn were from anemophilous prototypes. It is probably not possible to say that one of these conditions of itself is more primitive than the other, but the presence of one or the other may indicate relationships in particular cases.
[Go back carpel-call1-b]
The carpel is the basic foliar unit of the female reproductive organ of the angiosperm flower. It is an ovule-bearing structure evolved from what is currently and generally accepted to have been a nonphotosynthetic foliar leaflike appendage, and as such was a megasporophyll. This nongreen leaflike appendage is believed generally to have had a development parallel with that of the foliage leaf. It is not believed to have been derived from a foliage leaf. When solitary in the flower, the folded carpel and its stigmatic surface are better known taxonomically as the ovary and stigma. These two structures (ovary and stigma), plus the style (or styles) when present, comprise the pistil. The term carpel sometimes is used in taxonomic works in instances where the term pistil is the more common.25 (fn072-25) The terms are not always synonymous. The ovary of a pistil may be unicarpellate or it may have been formed by the
Lawrence 073
union of more than one carpel. Another term applied to the pistil is

 gynoecium, a collective term for the female element of the flower irrespective
of whether it is represented by one or by many pistils (Fig. 5 a-c).
gynoecium, a collective term for the female element of the flower irrespective
of whether it is represented by one or by many pistils (Fig. 5 a-c).
The gynoecium provides many taxonomic characters of major significance. In order to have a clearer comprehension of the importance, of the gynoecium and of the phyletic values ascribed to its components, it is desirable to know something of the morphological theories proposed to account for its origin and types. The most deeply entrenched and widely
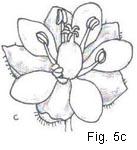
Fig.5. Gynoecial types
Fig. 5. Gynoecial types: a, pistils many and spiralled, the gynoecium apocarpous; b, pistils, many and whorled (cyclic). the gynoecium apocarpous; c, pistil solitary, the gynoecium syncarpous; d, pistil one, ovary compound; e, same, ovary vertical section; f, same, ovary cross-section; g, same, style apex and 3-lobed stigma; h, pistil one, ovary simple; i, same, vertical section; j, same, ovary cross-section; k, same, style-tip and simple stigma.
accepted theory for the origin of the carpel is the Candollean or appendicular theory. It was first proposed by Goethe (1790),26 (fn073-26) amplified by de Candolle (1827), revised and supported by van Tieghem (1871), and more recently defended on the basis of detailed anatomical studies by Eames (1931), Arber (1937), and others.27 (fn073-27) The fundamentals of this theory are that the antecedent of the carpel (and of which no living examples exist) was a nonphotosynthetic foliar appendage, probably a leaflike palmately 3-veined dorsiventral structure supplied with 3-vascular strands or traces. Functionally it is believed to have been originally
Lawrence 074
an open flat megasporophyll bearing ovules on its margins
(Fig. 6a),
 and to
have folded lengthwise with the ovules inside and margins fused (Fig. 6c). The
resultant structure is a unicarpellate, 1-loculed ovary. It is the product of a
single carpel, and is termed a simple ovary. Only one carpel (i.e., only one
foliar unit or megasporophyll) contributed to its
and to
have folded lengthwise with the ovules inside and margins fused (Fig. 6c). The
resultant structure is a unicarpellate, 1-loculed ovary. It is the product of a
single carpel, and is termed a simple ovary. Only one carpel (i.e., only one
foliar unit or megasporophyll) contributed to its
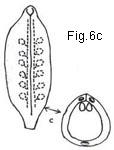
Fig.6. Hypothetical evolution of an ovary
Fig. 6. Hypothetical evolution of simple and compound ovary. a, three-lobed carpel with submarginal ovules; b, same, somewhat involute; c, simple ovary derived from "b" by infolding of ovules and connation of ventral margins; d, axis bearing three involute open carpels; e, compound ovary derived from "d" by connation of edges of adjoining carpels; f, axis with three open carpels with adjoining sides more or less parallel; g, compound ovary derived from "f" by connation of adjoining sides and margins; h, cross-section of "g" (hypothetical); i, cross-section of "g" (actual) showing loss of carpellary demarcation in the three septa. (Note: vascular strands shown with xylem elements blackened.)
formation. In the cross-sectional view of a simple ovary (Fig. 6c) note the locations of the midrib and the two marginal veins of the carpel, observing that the latter are in a relatively close proximity to one another and furthermore that the two rows of ovules (indicated diagrammatically in the expanded carpel) are so close together within the simple ovary as to appear almost as if in a single row. The zone or area occupied by this
Lawrence 075
row of ovules is the placenta. Examples of this type of ovary are to be found in the Leguminosae and many Ranunculaceae. Unlike the situation in animal anatomy, the angiosperm placenta is not a particular kind of tissue. It is the place where the ovules are attached.28 fn075-28) A simple ovary, inasmuch as it is the product of a single carpel, always has a single placenta. Furthermore, the single placenta of a simple ovary is parietal in position (i.e., it always is attached to the side of the locule wall, or to an intrusion of the side wall into the locule).29 (fn075-29)
In many plant families the gynoecium is composed of several to many simple pistils
(i.e., pistils each with a simple ovary) as in most of the Ranunculaceae. This
type of gynoecium is currently believed to represent a primitive phyletic
condition. It is from such a gynoecium, or from types ancestral to it, that the
compound ovary is believed to have developed. A compound ovary is any
ovary comprised of two or more carpels. If, perhaps by compression and fusion of
adjoining ovarian tissues, several simple pistils are connate (grown together),
the result is a single pistil of several carpels. Figure 6g-h is a diagram of the
ovary of such a pistil. Note that fusion is incomplete in
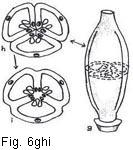 that the midrib and marginal veins of each of the 3 carpels are indicated. The placental
areas of each of the 3 original simple ovaries are shown to be converged about an
imaginary central axis. By the fusion of the 3 sets of walls of as many
adjoining simple ovaries, the resultant compound ovary is one of 3 locules, each
with a row of ovules in the axil formed by the septa (Fig. 6i). The placentation
of this type of compound ovary is said to be axile, since the ovules are
on the central axis which has been formed by the fusion of the carpel margins.30
(fn075-30)
Any ovary having axile placentation
that the midrib and marginal veins of each of the 3 carpels are indicated. The placental
areas of each of the 3 original simple ovaries are shown to be converged about an
imaginary central axis. By the fusion of the 3 sets of walls of as many
adjoining simple ovaries, the resultant compound ovary is one of 3 locules, each
with a row of ovules in the axil formed by the septa (Fig. 6i). The placentation
of this type of compound ovary is said to be axile, since the ovules are
on the central axis which has been formed by the fusion of the carpel margins.30
(fn075-30)
Any ovary having axile placentation
Lawrence 076
[UKT: In the original book, the whole of p076 was on Fig. 7. However, there was not enough place for all the explanations. I found these on p077 and they have been moved to p076 in this digitised version.]
Fig. 7. Presumed evolution of placentation types
Fig. 7. Presumed evolutionary development of ovary and placentation types. A: trilocular ovary with axile placentation, derived as shown in Fig. 6; Aa, schematic view of ovary; Ab, cross-section of same showing carpellary vascularization (xylem elements blackened). B: intermediate stage between "Ab" and "C" note partial loss of septa and retention of ventral vascular strands (each adjoining pair fused resulting in a reduction of six strands to three). C: unilocular ovary with free-central placentation, derived from "Aa"; Ca, schematic view of ovary; Cb, cross-section of same, differs from "B" only in complete loss of septation. D: compound ovary with basal placentation, derived from "Ca" by reduction of central placentae. E: compound ovary with single basal ovule, derived from "D" by ovule reduction. F: cross-section of unilocular tricarpellate ovary with parietal placentation derived from "Ab." G: same as "F" but septa reduced. H: advanced stage of "G" (no placental intrusion). I: hypothetical primitive situation of axis with three open carpels; la, schematic view; Ib, cross-section of same. J:compound ovary (unilocular, tricarpellate) with parietal placentation, derived by connation of adjoining carpel margins. K: compound ovary with parietal placentation, derived from "J"; Ka,
[UKT: Lawrence still had to explain about L to N.
This I found on p077 and have been moved to here below.]
schematic view of ovary; Kb, cross-section of same (note union of adjoining ventral carpellary strands and compare xylem orientation with that shown in "F," the latter presumed the more advanced condition). L: compound ovary with parietal placentation, derived from "K" (note absence of intruding carpellary margins); La, schematic view of ovary; Lb, cross-section of same (compare normal xylem orientation of ventral strands with reversed positions in "H," the retention of each pair of strands in "Lb" a less advanced situation than the fusion of them as shown in "H"). M: compound ovary with basal placentation, derived by placental reduction of "H" or of "La" N: compound ovary with single basal ovule, derived from "M" by ovule reduction.
Lawrence 077
tion (irrespective of the number of locules) is always a compound ovary because it is one that has been formed by the connation of two or more carpels. It is composed of as many carpels as it has placentae, and each carpel is usually represented by a locule.
A simple ovary typically has parietal placentation (Fig. 6c). This same apparent
placental type may occur in a compound ovary and by one of at least two
evolutionary lines of development. Presumably it could have
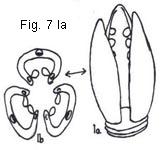 arisen from a gynoecium of open and incurved carpels standing in close proximity to one
another and which became fused together marginally. Figure 7Ia indicates diagramatically such a situation involving 3 carpels. A single pistil results
from the marginal connation of these carpels. It is a pistil that has a compound
ovary of 1 locule and 3 parietal placenta. Note that each placental area is
formed from the union of two adjacent carpel margins, each bearing a row of
ovules and that the midrib of each carpel stands about midway between the
placentae. The second line of evolutionary development believed responsible for
some examples of parietal placentation in multicarpellate ovaries is that
derived from an ovary having axile placentation. Figure 7A-7H illustrates this
situation. In this hypothetical instance, a 3-carpelled and 3-loculed ovary
gives rise to a 3-carpelled unilocular ovary by separation of the 3 central
placental zones and their recession to or toward the ovary walls. The number of
carpels remains the same, but the placental position is changed from axile to
parietal, and when the 3 locules open into each other a single locule results.
It is usually true that an ovary with parietal placentation, irrespective of the
number of carpels, is an ovary of a single locule.31
(fn077-31)
In many compound ovaries of this type, the placentae are not restricted to the
inner periphery of the ovary wall but are on intrusions of the ovary wall (Fig.
7G) or (as in Coptis or Gentiana spp.) are on the
inward-projecting free margins of the carpel or of adjoining carpels. By some
authors this placental condition is termed falsely parietal, but macroscopic
arisen from a gynoecium of open and incurved carpels standing in close proximity to one
another and which became fused together marginally. Figure 7Ia indicates diagramatically such a situation involving 3 carpels. A single pistil results
from the marginal connation of these carpels. It is a pistil that has a compound
ovary of 1 locule and 3 parietal placenta. Note that each placental area is
formed from the union of two adjacent carpel margins, each bearing a row of
ovules and that the midrib of each carpel stands about midway between the
placentae. The second line of evolutionary development believed responsible for
some examples of parietal placentation in multicarpellate ovaries is that
derived from an ovary having axile placentation. Figure 7A-7H illustrates this
situation. In this hypothetical instance, a 3-carpelled and 3-loculed ovary
gives rise to a 3-carpelled unilocular ovary by separation of the 3 central
placental zones and their recession to or toward the ovary walls. The number of
carpels remains the same, but the placental position is changed from axile to
parietal, and when the 3 locules open into each other a single locule results.
It is usually true that an ovary with parietal placentation, irrespective of the
number of carpels, is an ovary of a single locule.31
(fn077-31)
In many compound ovaries of this type, the placentae are not restricted to the
inner periphery of the ovary wall but are on intrusions of the ovary wall (Fig.
7G) or (as in Coptis or Gentiana spp.) are on the
inward-projecting free margins of the carpel or of adjoining carpels. By some
authors this placental condition is termed falsely parietal, but macroscopic
[UKT: explanations found in this place has been moved to p076]
Lawrence 078
examination of a given ovary seldom affords a means of determining clearly between truly parietal and falsely parietal. The distinction is of morphological and phyletic significance, but it of little taxonomic use in ordinary identification procedures.
A third basic type of placentation within a compound ovary is that known as free-central placentation (see Fig. 7Ca). It is generally agreed that this type has been evolved from the axile type of placentation by the persistence of the central column and its placentae and the disappearance of the partitions, or septa. There is evidence that, in a few genera having free-central placentation (e.g., Primula), the axis on which the ovules are borne is comprised of carpellary tissue into which extends stem tissue for a short distance, the latter representing the extension of the stele from the subtending pedicel. An ovary with free-central placentation always has a single locule, but is composed of two or more carpels and hence is always a compound ovary.32 (fn078-32) However, since the placental zones are closely appressed laterally about the axis and usually without any lines of demarcation between them, it is necessary to resort to other means, as indicated below, to determine or approximate the carpellary number.
The reduction of the central axis in the free-central type of placentation to a nubbin terminated by a single ovule produces a type designated as basal placentation, and taxonomically this comprises a fourth type of placentation. 33(fn078-33) In its extreme form, none of the central axis remains and the ovule appears as if on the floor of the locule. In other instances, the situation where the number of apparently basal ovules is more than one (usually 2 to 4) represents a condition believed to have been derived from axile placentation types or, in unilocular ovaries this probably has come through the free-central stage (cf. Figs. 7D and 7E). Taxonomists, in descriptions and keys, generally refer to them as basal when no axile condition is macroscopically apparent. A uniovulate placentation in some instances has been derived from parietal placentation of a multicarpellate ovary (Gramineae, Compositae) or of a unicarpellate ovary (as in some Ranunculaceae or Rosaceae). Because of these two derivations of a uniloculate 1-ovuled ovary, it is apparent that although the uniovulate
Lawrence 079
ovary appears to be simple, it may represent a highly advanced condition and
may be a simple or a compound ovary (cf. Fig. 7N). The pendulous ovule,
characteristic of some uniovulate ovaries, usually represents a reduction from
parietal placentation but may have been derived from axile placentation (as in
some Caprifoliaceae).
Taxonomically, these types of placentation provide characters of considerable
significance and utility. Very often they are indicators of phyletic
relationship between and within major groups. They are characters that
frequently are employed in analytical keys, especially in keys to families. It
is important to be able to differentiate and recognize the different types, and
to have an intelligent concept as to their relative level of development from
the primitive simple ovary with its parietal (or marginal) placentation, to the
extreme degree of reduction represented by the uniloculate 1-ovuled simple or
compound ovary. Occasionally it is desirable to know the number of carpels
represented in an ovary. Accurate determination of carpellary number frequently
requires microscopic study of a series of microtome sections, and in many
instances may even then be determined only by comparative studies. However, the
number of styles, stigmas, or stigma branches or lobes of a pistil often serve
as indicators of the number of carpels present. In general, if the number of any
one of these features is two or more, it may be assumed that the ovary bearing
them is a compound rather than a simple ovary. An ovary of two or more complete
locules is always compound. An ovary of a single locule is simple only
if it has a single parietal zone of placentation and is comprised of a single
carpel.
The position of an ovary is referred to as
being superior,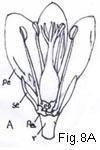 inferior, or half inferior. A superior ovary is one that
is situated above the point of attachment of perianth and androecium. An ovary
whose position is superior is considered to represent the
inferior, or half inferior. A superior ovary is one that
is situated above the point of attachment of perianth and androecium. An ovary
whose position is superior is considered to represent the
 primitive position. An
inferior ovary is one situated below the apparent point of attachment of
the perianth and androecium. An ovary whose position is half inferior
(or subinferior) is more or less intermediate between these two. (For diagrams
of these types, see Fig 8.)
primitive position. An
inferior ovary is one situated below the apparent point of attachment of
the perianth and androecium. An ovary whose position is half inferior
(or subinferior) is more or less intermediate between these two. (For diagrams
of these types, see Fig 8.)
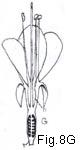
[UKT: Fig.8A - superior ovary; Fig.8G - inferior ovary.]
There are two theories to account for the evolution of the inferior from the superior ovary, and in some instances the situation is explained only by accepting views from each of them. In a typical angiosperm flower the 5 whorls or series of components (calyx, corolla, two whorls of stamens, and the gynoecium) are borne on an axis called the receptacle or torus (Fig. 8A). The older and perhaps until recently the more widely accepted theory proposed to account for the derivation of the inferior ovary from
Lawrence 080
the superior is the receptacular theory, by which it was concluded that the inferior ovary is embedded in or surrounded by a tube or cup of receptacular tissue. By this theory it was held also that in those flowers whose ovary is superior, but surrounded by a cuplike or cylinderlike tube called the hypanthium (more commonly and inaccurately termed the calyx tube and by some morphologists called the floral tube) (Fig. 8B and F), the hypanthium itself is of receptacular tissue (Fig. 8Ca). Anatomical studies of a wide range of materials by Eames (1931), Jackson (1934), and MacDaniels (1937) indicated that, for the majority of
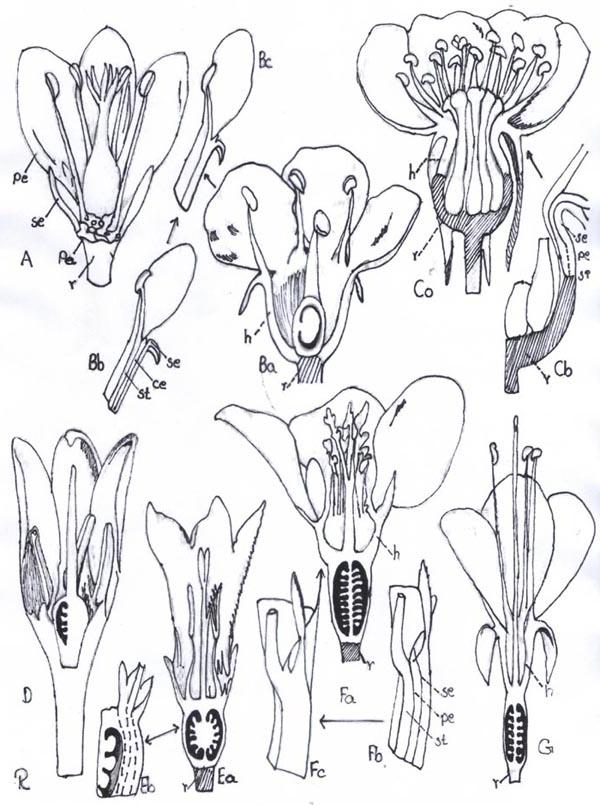
Fig. 8. Ovary position. A, ovary superior, perianth and stamens hypogynous; B, ovary superior, perianth segments and stamens perigynous; Ba, vertical section; Bb, detail of Ba to show composition of hypanthium: Bc, same, minus lines of adnation of perianth and androecium; C, ovary superior, perianth and stamens perigynous, hypanthium of "Rosa" type; Ca, vertical section; Cb, detail to show relation of receptacular cup to hypanthium and composition of latter: D, ovary half-inferior, stamens perigynous; E, ovary inferior, perianth and stamens epigynous; Ea, vertical section of flower; Eb, detail of same to show adnation of hypanthium to ovary; F, ovary inferior, hypanthium present, perianth and stamens epigynous; Fa, vertical section of flower; Fb, detail to show components of hypanthium, Fc, section of hypanthium; G, ovary inferior, stamens exserted, hypanthium present. (h hypanthium, pe petal, r receptacle, se sepal, st stamen.)
Lawrence 081
flower possessing inferior ovaries, or ovaries within an hypanthium, the tissues previously believed to have been receptacular in origin are formed form the fusion of the foliar or appendicular units as represented by perianth parts and stamens (Fig. 8E). This fusion represents a phylogenetic, and not an ontogenetic, phenomenon.
The second, and more widely accepted, of the two theories is known as the appendicular theory. By this view, it is held that the hypanthium that may surround the superior ovary, and the tissues that are adnate to of the flower (i.e., they are appendicualr in origin) and are not of the axis, receptacle, or torus. In most instances (an exception being the Santalaceae and Calycanthaceae), no stelar tissues are found in the hypanthium, irrespestive of whether it be free or adnate to the ovary. In the genus Rosa it has been established that only the upper part of the hypanthium is appendicular in origin, for it was shown by Jackson (1934) that the basal half or less has resulted from the peripheral extrusion of the apex of the apex of the floral axis (receptacle).34 (fn081-34) There is ample evidence that the inferior ovary of present-day plants represents two lines of evolutionary development: (1) the usual situation represented by adnation of foliar elements (calyx, corolla, androecium) to the ovary wall and (2) the uncommon condition represented by adepression and peripheral extrusion of the receptacle with the latter surrounding and adnate to the ovary. On the basis of recent studies, made independently by several workers, it is now generally accepted that while the inferior ovary represents an advanced condition derived form the superior ovary, "epigynous and perigynous flowers differ in no essential respect form the hypogynous condition; the inferior ovary is, in most cases at least, appendicular in origin, not receptacular," (Wilson and Just, p. 103).
Most phylogenists have placed much emphasis on the phyletic significance of ovary position, and to the extent that families have been classified in orders largely on the basis of this character. It is also true that the orders composed of families that are characterized by both types of ovary position have been grouped together in most classifications. In general, the ovary position provides a character of considerable reliability, although intergradation occurs between all three ovary position types in a few genera and families.
fn065-19. By "dorsiventral compression and lateral fusion
of photosynthetic branches" is meant the flattening of presumably green terete
secondary stems in a single plane with their posterior (inner) surfaces in a
ventral (uppermost) position, and furthermore, that by marginal connation of
adjoining edges a green leaflike organ evolved. By this view, any leaf is a
derivative of a group of branches and because of this origin represents a highly
modified branch system.
Go back fn065-19b
fn065-20. The views presented here to account for the
nature and origin of the angiospermous flower are believed to represent opinions
of the majority. However, there are other and contradictory views on these
matters, and for an introduction to them the reader is referred to the papers by
Arber (1937), Bancroft (1935), and Ozenda (1949), wherein the opposing views are
reviewed but not supported. Of these, perhaps the most widely publicized was
Saunders' theory of carpel polymorphism, early refuted by Eames
(1931) and currently rejected by most investigators.
Go back fn065-20b
fn067-21. See footnote 38 (p.84) for explanation of the
terms distinct and connate.
Go back fn067-21b
fn067-22. Many taxonomists assume the pappus of the
Compositae flower to represent in all genera a reduced and modified calyx.
Studies by Koch (1930) of the floral anatomy in the form the corolla of some
present-day members. In those members of the family where this has been
suggested the pappus is presumed to be bracteate in origin. This latter view
also provides plausible explanation for the origin of the apparent double pappus
encountered in many genera of the family, in which situations the inner one may
or may not be a modified calyx and the outer one perhaps derived by modification
of bracts.
Go back fn067-22b
fn070-23. Corner (1946) cited the following families as
characterized in part by having centrifugal androecia: Actinidiaceae. Aizoaceae,
Bixaceae, Cactaceae, Capparidaceae. Dilleniaceae, Hypericaceae, Loasaceae,
Lecythidaceae, Malvaceae, Paeoniaceae, Theaceae, Tiliaceae.
Go back fn070-23b
fn071-24. Viable fresh pollen grains (of nonhybrid
origin) contain an abundance of chromatin material. Smears or smashes of such
pollen are stained readily with aceto-carmine solution or aniline (cotton) blue,
and an indication of possible hybridity may be demonstrated if the stained
smears of pollen of suspected hybrids fail to take on any of the stain.
The technique and interpretation of the findings require skill and experience. A
magnification of X 120 is generally adequate in the application of this
preliminary test. It is emphasized that this test may only indicate possible hybridity and results should never be interpreted as presumptive of hybridity.
Go back fn071-24b
fn072-25. There is no uninamity of opinion concerning the
definition of the term pistil. Some taxonomists, and many morphologists,
consider the term a source of confusion and abandon it. In its place they apply
the term carpel to all unicarpellate organs and gynoecium to all multicarpellate
female elements within a single flower (the latter use, irrespective of the
number of free ovaries present). Basis for this view lies in the contention that
the term pistil has been applied variously during the years of descriptive
botany. These reasons for rejection of the term pistil are considered untenable
and use of the term is defended, on the grounds that to a taxonomist (working
primarily with macroscopic organs) 3 basic terms are needed to discuss the
essential features of the female element: (1) the megasporophyll or foliar
element, (2) the gross structure represented by a single ovary with its style
(or style) and stigma (or stigmas), irrespective of the number of foliar
elements entering into its composition and of the freedom of fusion of those
elements, and (3) the female element or household of the flower in toto,
irrespective of the number of free and distinct ovaries involved. The 3 terms
that meet these taxonomic needs are, respectively: carpel, pistil and gynoecium.
As defined above, the term pistil is neither ambiguous nor confusing. It is used
by current taxonomic workers and was used (among others) by Linnaeus, Ludwig,
Mirbel, St. Hilaire, de Candolle, Robert Brown, Hooker, and Asa Gray.
Go back fn072-25b
fn073-26. By Goethe's concept, the primitive carpel was
held to be a green photosynthetic leaf bearing marginal ovules, a view not now
accepted.
Go back fn073-26b
fn073-27. Three current theories held in opposition to
this view are to the effect that: (1) carpels have originated from the floral
axis and are axillary organs, (2) all floral organs are structures that always
have existed, as organs sui generis, and (3) carpels do not exist at all
(acarpy), a theory advocated by Thompson.
Go back fn073-27b
fn075-28. In some plants (as the tomato, Lycopersicon, or
in Epigaea) the cells of the placental area proliferate and form a prominent
tissue mass that extrudes into the locule and is properly referred to as the
placenta or placental zone. See Fig. 273.
Go back fn075-28b
fn075-29. Some botanists designate this placental
condition as marginal or ventral, pointing out correctly that it differs
from the parietal placentation of a syncarpous (i.e., compound) ovary in the
placenta representing two connate margins of the same carpel and not the connate
margins of two adjacent carpels. This distinction is phyletically important but
is not always apparent from macroscopic examination common to taxonomic study
(for example, only by knowledge of floral anatomy or comparative morphology can
one identify a unilocular ovary with a single placenta and seemingly with
marginal placentation, to be a tricarpellate ovary derived by reduction from an
ancestral form that was trilacular with axile placentation). Parietal is
accepted here as a descriptive term, applicable to any placenta situated on the
ovary wall or an intrusion of that wall.
Go back fn075-29b
fn075-30. Axile placentation is the result of the union
of carpel margins at the center of the ovary. The term axile indicates merely
that the ovules are situated in the axil of two adjoining septa. It should be
understood clearly that in axile placentation no true axis or stem is present in
the central column, that there is no stelar tissue involved (in Pyrus and
Cydonia the stem tip may extend a short distance up between the carpels).
It is believed that the ovules of all angiospermous overies are always of foliar
and never of cauline origin.
Go back fn075-30b
fn077-31. One exception to this occurs in the ovary of
members of the family Cruciferae, where the ovary is normally 2-loculed and
4-carpelled with the ovules attached parietally along one or both peripheral
margins of the commissural sides of two of the carpels. (See Fig. 152
Go back fn077-31b
fn078-32. The central column consists of a placental mass
of proliferated tissue composed of as many fused placentae as there are carpels.
The number of placentae contributing to this mass is usually not evident, but is
strikingly so in some genera of Caryophyllaceae and in a few genera of
Primulaceae.
Go back fn078-32b
fn078-33. A fifth type is that known as lamellate
placentation, occurring when the placental area and its ovules covers one or
more broad, flattened, and usually lamellate surfaces which may be peripheral or
represent platelike septa (as in Papaver, Nuphar, Nymphaea,
and Butomus).
Go back fn078-33b
fn081-34. For a review of the pertinent literature, see
Wilson and Just (1939) and Douglas (1944)
Go back fn081-34b
(in alphabetical order)
acropetal, basipetal, centripetal
-petal suff. 1. Moving toward: basipetal. [From New Latin -petus from Latin petere to seek; See pet- in Indo-European Roots.] -- AHTD
acropetal adj. 1. Developing or maturing from the base toward the apex, as in those plant organs in which the younger tissues are nearer the apex. -- AHTD
basipetal adj. Botany 1. Of or relating to the development or maturation of tissues or organs or the movement of substances, such as hormones, from the apex downward toward the base. -- AHTD
centripetal adj. 4. Botany Developing or progressing inward toward the center or axis, as in the head of a sunflower, in which the oldest flowers are near the edge and the youngest flowers are in the center. -- AHTD
Go back acropetal-b, basipetal-b, centripetal-b
adnate adj. Biology 1. United to a part or organ of a different kind, as stamens attached to petals. [Latin adnātus variant of agnātus, past participle of agnāscīto grow upon; See agnate ] -- AHTD
Go back adnate-b
andro- or andr- pref. 1. Male; masculine: androgen. 2. Stamen or anther: androecium. [Greek from anēr andr-man; See ner- 2 in Indo-European Roots.] -- AHTD
androecium n. pl. androecia 1. The stamens of a flower considered as a group. -- AHTD
Go back androecium-bgyno or gyn- pref. 1. Woman: gynarchy. 2. Female reproductive organ; pistil: gynophore. [From Greek gunē woman; See g w en- in Indo-European Roots.] -- AHTD
gynoecium n. pl. gynoecia 1. The female reproductive organs of a flower; the pistil or pistils considered as a group.-- AHTD
Go back androe-gynoe-cium-b
anthesis n. Botany 1. The period during which a flower is fully open and functional. [Greek anthēsis flowering from anthein to bloom from anthos flower]
Go back anthesis-b
floral envelope n. 1. The perianth of a flower. -- AHTD
Go back flo-envel-b
nubbin - 1. a small stunted ear of corn. 2. A small stunted or projecting part -- www.thefreedictionary.com/nubbin
Go back to nubbin-b
peri- pref. 1. Around; about; enclosing: perimysium. 2. Near: perinatal. [Greek from peri around, near; See per 1in Indo-European Roots.] -- AHTD
perigone - collective term for the outer parts of a flower
consisting of the calyx and corolla and enclosing the stamens and pistils.
syn. chlamys, floral envelope, perianth, perigonium. --
www.thefreedictionary.com/perigone
Go back perigone-b
peri- pref. 1. Around; about; enclosing: perimysium. 2. Near: perinatal. [Greek from peri around, near; See per 1 in Indo-European Roots.] -- AHTD
Go back peri-b
pistil n. 1. The female, ovule-bearing organ of a flower, including the stigma, style, and ovary. [French from New Latin pistillum from Latin pestle (from its shape)] -- AHTD
There is no unanimity of opinion concerning the definition of the term pistil. -- fn072-25
Go back pistil-call1-b pistil-call2-b pistil-call3-b
sporangium n. pl. sporangia 1. A single-celled or many-celled structure in which spores are produced, as in fungi, algae, mosses, and ferns. Also Called spore case . [New Latin spor(o)- Greek angeion vessel; See angio- ] -- AHTD
Go back sporangium-b
n. pl. steles or stelae 1. An upright stone or slab with an inscribed or sculptured surface, used as a monument or as a commemorative tablet in the face of a building. 2. Botany The central core of vascular tissue in a plant stem or root. [Greek stēlē pillar; See stel- in Indo-European Roots.] stelar adj. -- AHTDstele
Go back stele-b
theca
n. pl. thecae 1. A case, covering, or sheath, such as the pollen sac of an anther, the spore case of a moss, or the outer covering of the pupa of certain insects. [Latin case, receptacle from Greek thēkē] -- AHTDUKT: Compare 'theca' to Burmese-Myanmar
![]() {thét~ta} meaning 'box'. Caveat: this observation is mine, not that of a
linguistic authority. At this time in life (I am 72), I am in transliterating
Burmese-Myanmar to English-Latin.
{thét~ta} meaning 'box'. Caveat: this observation is mine, not that of a
linguistic authority. At this time in life (I am 72), I am in transliterating
Burmese-Myanmar to English-Latin.
Go back theca-b
tetramerous adj. 1. Having or consisting of four similar parts. 2. Botany Having flower parts, such as sepals, petals, and stamens, in sets of four, as in the evening primrose. -- AHTD
Go back tetramerous-b
End of TIL file