
flower-types.htm
by George H. M. Lawrence, Professor of Botany at the Bailey Hortorium, Cornell University, 1951
Photo-copied by Maung Kan Tun from the original text.
Edited and set in HTML by U Kyaw Tun (M.S., I.P.S.T., USA),
Tun Institute of Learning (TIL) and the staff of TIL.
Not for sale. No copyright. Free for everyone.
Prepared for students and staff of TIL
Computing and Language Center, Yangon, MYANMAR :
http://www.tuninst.net ,
http://www.softguide.net.mm ,
www.romabama.blogspot.com
index.htm |
Top
Lawrence-indx.htm
Principles of Taxonomy
Flower types
Flower types p081
Fig. 9. Flower types and arrangement p082
Distribution of
sexes p.082
Disposition of
perianth parts p.084
Fusion
and modification of perianth parts p.084
Fruits p.085
Seeds p.087
Embryology p.087
Karyology p.088
Literature p.088-091
Footnotes
UKT notes
[{cont. p081}]
Modern taxonomy demands an understanding of the morphological composition of the
flower. Descriptive taxonomic literature [{p082}]
is predicted on an understanding of the various types of flowers as indicated by
gross morphology, and a knowledge of flower types is essential to satisfactory
use of keys and descriptions available in floras and manuals of vascular plants.
Many of the floral conditions indicated below are of little phyletic
significance, and often are less reliable than is indicated by their extensive
use by many authors. In addition to
knowing these floral types, one should endeavor to grasp some comprehension of
their relative constancy and reliability for use in the characterization or
differentiation of taxonomic units.
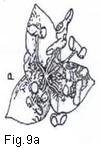
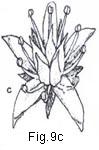
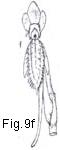
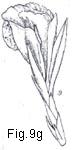
Fig. 9. Flower types and arrangement
Fig. 9. Flower types and arrangement: a, flower staminate (perianth uniseriate); b, flower pistillate (perianth uniseriate); c. flower bisexual (perianth biseriate and corolla polypetalous); d, flowers unisexual (plant monoecious); e, flower actinomorphic or regular; f, flower zygomorphic; g, flower irregular; h, flower gamopetalous.
A flower represented by the two perianth series, the androecium, and gynoecium,
is a complete flower; one lacking the perianth series but possessing both
sex elements is an incomplete flower, but since it possess both kinds of
sex elements it is termed a [{p083}]
 perfect flower and (with or without the perianth) is often referred to as
a bisexual or hermaphroditic. A flower lacking the organs of one
sex is an imperfect flower and is unisexual (Fig. 9a, b).
Irrespective of the perianth situation, a unisexual
perfect flower and (with or without the perianth) is often referred to as
a bisexual or hermaphroditic. A flower lacking the organs of one
sex is an imperfect flower and is unisexual (Fig. 9a, b).
Irrespective of the perianth situation, a unisexual
 flower possessing only an
androecium is a staminate flower35 (fn083-35),
while one possessing only a gynoecium is a pistillate flower.
flower possessing only an
androecium is a staminate flower35 (fn083-35),
while one possessing only a gynoecium is a pistillate flower.
[UKT:Fig. 9a - flower staminate (perianth uniseriate);
9b - flower pistillate (perianth uniseriate)]
Some species of plants characterized by unisexual flowers have the staminate
flowers restricted to one plant and the pistillate flowers restricted to
another. Such plants are dioecious (e.g., Salix, Myrica,
Garrya, most spp. of Fraxinus). In other unisexually flowered taxa the
pistillate flowers and the staminate flowers are on the same plant
 (Fig. 9d).
Such a plants is monoecious (e.g., Zea, Betual, Pinus,
Euphorbia).36 (fn083-36)
Some plants are characterized by the presence of both bisexual and unisexual
flowers in the same inflorescence or on different parts of the plant. Such
inflorescences or plants are said to be polygamous (e.g., many members of
the Compositae). A derived condition is that existing when a plant is
functionally dioecious, but has scattered throughout its inflorescences a few
perfect flowers (e.g., some spp. of Ilex, Celastrus, Acer,
Rhus). Such plants are said to be polygamodioecious.
(Fig. 9d).
Such a plants is monoecious (e.g., Zea, Betual, Pinus,
Euphorbia).36 (fn083-36)
Some plants are characterized by the presence of both bisexual and unisexual
flowers in the same inflorescence or on different parts of the plant. Such
inflorescences or plants are said to be polygamous (e.g., many members of
the Compositae). A derived condition is that existing when a plant is
functionally dioecious, but has scattered throughout its inflorescences a few
perfect flowers (e.g., some spp. of Ilex, Celastrus, Acer,
Rhus). Such plants are said to be polygamodioecious.
The primitive type of flower is postulated by most modern morphologists to have been a complete flower, that is, it was composed of a perianth, androecium, and gynoecium. Any situation where one of these floral components is missing is considered to represent a derived or advanced condition. For example, a case of phyletic advancement is indicated by the suppression or reduction of parts, as when stamens are represented by staminodes (which may be nectiferous, petaliferous, or filamentous) or when the gynoecial elements are represented by pistillodes. The reduction may have taken place within all of the androecial elements (in which case the flower is unisexual and pistillate) or to have affected some or all of the carpellary elements of the gynoecium. In a few cases, both sex organs have become suppressed or reduced to a point of being wholly nonexistent or at least nonfunctional, with the floral axis terminated only by the perianth parts. Such a structure morphologically is not a flower, [{p084}] since it contains no reproductive elements, but in descriptive taxonomic literature is termed a neutral or sterile flower (e.g., some of the flowers of some spp. of Hydrangea, Viburnum, or Compositae).
 The arrangement of perianth parts in a spiral and
without sharp demarcation between calyx and corolla usually is considered the
most primitive (e.g., Magnolia, Nymphaea, Nelumbo). If one looks down on
an open flower of this type, one notes a symmetry of perianth parts, and it is
apparent that such a perianth can be bisected in two or more planes and in into
similar halves (as, for example, one can cut a pie into two equal halves from
any selected point of the circumference). Such a perianth is actinomorphic
or regular. 37 (fn084-37)
Flowers with perianth
The arrangement of perianth parts in a spiral and
without sharp demarcation between calyx and corolla usually is considered the
most primitive (e.g., Magnolia, Nymphaea, Nelumbo). If one looks down on
an open flower of this type, one notes a symmetry of perianth parts, and it is
apparent that such a perianth can be bisected in two or more planes and in into
similar halves (as, for example, one can cut a pie into two equal halves from
any selected point of the circumference). Such a perianth is actinomorphic
or regular. 37 (fn084-37)
Flowers with perianth
 series in whorls rather than spirals may, with equal
precision of application of the terms, have actinomorphic or regular perianths
(Figs. 9c, 9e). Sometimes the corolla may be actinomorphic and the calyx not so,
or the reciprocal condition may be present. In other plants the perianth parts,
or segments, are so
series in whorls rather than spirals may, with equal
precision of application of the terms, have actinomorphic or regular perianths
(Figs. 9c, 9e). Sometimes the corolla may be actinomorphic and the calyx not so,
or the reciprocal condition may be present. In other plants the perianth parts,
or segments, are so
 disposed that the floral envelopes may be bisected only
along
disposed that the floral envelopes may be bisected only
along
 one diameter or in one plane (e.g., in Orchidaceae, papilionaceous
Leguminosae, most Labiatae). These perianths are zygomorphic (Fig. 9f). In a few
groups the perianth, or its parts, is of such disposition that it is not
possible to bisect it into two like parts, and it is said then to be irregular
(Fig. 9g). Generally speaking, the term irregular is more often applicable to
flowers than to perianths (e.g., in Canna or Zingiberaceae).
one diameter or in one plane (e.g., in Orchidaceae, papilionaceous
Leguminosae, most Labiatae). These perianths are zygomorphic (Fig. 9f). In a few
groups the perianth, or its parts, is of such disposition that it is not
possible to bisect it into two like parts, and it is said then to be irregular
(Fig. 9g). Generally speaking, the term irregular is more often applicable to
flowers than to perianths (e.g., in Canna or Zingiberaceae).
Fusion and modification of perianth parts
The primitive situation of perianth parts has until recently been accepted to be that of distinct and free 38 (fn084-38) from one another. Most contemporary systems of angiosperm classification accept plants with perianth parts free and distinct to be more primitive than those with perianth parts of each series [{p085}] connate. Corollas composed of distinct parts are polypetalous (choripetalous, apopetalous) (Fig. 9c), those whose petals are to any degree marginally connate are gamopetalous (sympetalous) (Fig.9h). The gamopetalous condition may be very obvious (e.g., in Aster, Convolvulus, Digitalis) or superficially not present (e.g., in Armeria, Chionanthus) and in cases of deeply segmented corollas the condition may be determined only after careful dissection and study.
In general, most botanists accept the view that a perianth composed of free and distinct parts (as of sepals, petals or of tepals) is more primitive than one in which such parts are partially or completely connate or adnate (thus resulting in synsepally, sympetally, or the production of an hypanthium). There is abundant evidence indicating that taxa whose flowers exhibit fusion of perianth parts arose independently (i.e., by polyphyletic origins) in widely separated families and orders. A majority of morphologists, and many botanists, currently reject those phyletic views that would group all dicot orders whose families possess gamopetalous corollas (as, for example, the taxon Sympetalae). There is evidence also that some corollas now composed of distinct petals, or of segments connate only at the base, have been derived from ancestors whose corollas were conspicuously or completely gamopetalous (as in some Onagraceae, Plumbaginaceae). Likewise the vascular anatomy of some of these seemingly near-polypetalous corollas shows that their segments contain marginal vascular strands that clearly once belonged to adjoining lobes or segments of the same perianth series; this is evidence that in such situations the corolla segmentation probably was preceded by a more completely gamopetalous condition. Perianths having these complex vascular situations are believed generally to be more advanced phyletically than are those exhibiting simple but complete connation of parts. For these reasons it should be borne in mind that while the absence of fusion of perianth parts is generally to be construed to represent a primitive situation, it may in some taxa represent a very advanced phyletic situation.
A fruit may be defined as the product of the ripened ovary or pistal of a flower and may be composed in part also of accessory floral or vegetative parts. It is the seed-bearing (or containing) organ of a plant, and in this sense the ripened female strobilus (or cone) of a conifer may be referred to as a fruit. Fruits are important in the classification, delimitation, and identification of seed plants because generally they provide characters very reliable in the characterization of genera and of families. Many kinds and types of fruits exist. An appreciable [{p086}] number of classifications have been proposed for their systematic organization 39 (fn086-39); all are empirical and artificial. None has been devised during much of the last century, and none is based on the fundamental morphology of the structures concerned. The basic weakness of these classifications is their artificiality, and since they have been devised without due regard for the morphology of the ovaries and gynoecia from which the fruit is produced, the definitions of their categories or types invariably are violated by an abundance of exceptions. Until a more satisfactory classification has been devised, contemporary taxonomists are obliged to continue to employ the currently used conventional classification that follows the basic schema presented in Asa Gray's 6th edition of Structural botany or in Gray's lessons in botany. A synopsis adapted from that published by Gray (1879) and accounting for the more commonly encountered fruit types is given below. In making reference to it, the omnipresence of exceptions must be acknowledged.
[UKT: The synopsis (Fruits simple, the product of a single pistil)
referred to from the scanned page is given below. Definitions of fruits from the
synopsis is given in my notes.
Fleshy fruits: Berry,
Drupe, Hesperidium, Pepo, Pome
Dry fruits: Achene;
Follicle; Legume; Loment; Nut; Pyxis; Samara; Schizocarp; Utricle]


[{p087}]
Seeds are fertilized mature ovules and each contains an embryo. An abundance of
taxonomic and phylogenetic characters are provided by the seeds of plants.
Generally speaking, those of general taxonomic use are the more superficial and
associated with the seed coat (usually derived from the outer integument of the
ovule, sometimes termed testa). Outgrowths such as wings (as in the pine,
trumpet vine, or maple) or the coma (a tuft of hair as in milkweed, cotton) are
examples of this character. In other groups, the presence and character of pits,
sutures, sculpturing, and surface configurations are of significant value
taxonomically (Fig.322). Of more fundamental classifactory value are the
internal characters afforded by seeds of many plant groups; particularly is this
true of the presence or absence of endosperm (albumen), shape and position of
the embryo, and the character, number, and arrangement of the cotyledons. In
this regard, Martin (1946) devised a phylogenetic schema of seed plants based on
the comparative internal morphology of seeds that compares very favorably with
modern concepts based on combinations of other structures.
The taxonomic value of embryological data has been ignored by the systematist until recently, or has been considered useful only to the phylogenist dealing with the classification of taxa in the higher categories. In this regard it has been emphasized by Maheshwari (1945, 1950) and by Just (1946) that the taxonomist should be cognizant of the significance of these data and recognize their value and limitations. Embryological characters found to be of taxonomic and phyletic significance include those of the male and female gametophyte as well as of the embryo. Just (1946) proposed a set of symbols whereby the attributes of each of the pertinent characters might be designated in condensed form. In focusing attention on the use of these data by the taxonomist he stated (p.354):
The accepted orders of flowering plants may well be delimited differently, if embryological data are applied consistently, and groups which have so far not been investigated are studied embryologically. Therefore, the use of embryological data will be greatest as supplementary evidence in the improvement of our systems of [{p088}] classification, particularly with regard to the determination of the correct position and affinities of families and orders, whereas their application in purely descriptive works such as floras and manuals as well as in teaching elementary plant taxonomy may never prove feasible.
Embryological data need not be accorded more recognition than other taxonomically valuable characters. They do, however, deserve their rightful place among the others, a position they have not yet attained in the eyes of all botanists.
Much earlier, Rutgers (1923) proposed the use of formulae by which to designate embryo-sac and embryo conditions, but made no attempt to include the range of gametophytic characters of both sexes that were taken into account by Just. The recent contribution by Johansen (1950) should do much to enlighten the taxonomist of the value of embryological data, and provides a ready and classified reference to these data and to the pertinent literature.
Cytology and cytogenetics have provided many data concerning the number, morphology, and behaviour of chromosomes within the nucleus of the gametophyte that are of taxonomic significance. For a discussion of this see Chapter 8.
Anderson, E. The concept of the genus. II. A survey of modern opinion. Bull. Torrey Bot. Club, 67: 363-369, 1940.
Arber, A. The interpretation of the flower: a study of some aspects of morphological thought. Biol. Rev. 12: 157-184, 1937.
Babcock, E. B. Systematics, cytogenetics and evolution in Crepis. Bot. Rev. 8: 139-190, 1942.
Bancroft, H. A review of researches concerning floral morphology. Bot. Rev. 1: 77-99, 1935.
Bartlett, H. H. The concept of the genus. I. History of the generic concept in botany. Bull. Torrey Bot. Club, 67: 349-362, 1940
Briquet, J., ed. International rules of botanical nomenclature. ed. 3, Jena, 1935.
Brown, W. H. The bearing of nectaries on the phylogeny of the flowering plant. Proc. Amer. Philosoph. Soc. 79: 549-594, 1938.
Buchholz, J. T. Seeds. Sci. Monthly, 38: 367-369, 1934.
Camp, W. H. The concept of the genus. IV. Our changing generic concepts. Bull. Torrey Bot. Club, 67: 381-389, 1940.
Camp, W. H. and Gilly, C. L. Floral abnormalities in Linaria vulgaris, with
notes on a method by which new genera may arise. Torreya, 41: 33-42, 1941.
_____. The structure and origin of species. Brittonia, 4: 325-385, 1943.
Celakovsky, L. F. Gedanken iiber eine zeitgemässe Reform der Theorie der Blütenstände. Bot. Jahrb. Engler, 16: 33-51, 1892.
Chute, H. M. The morphology and anatomy of the achene. Amer. Journ. Bot. 17: 703-723, 1930.
Clausen, J., Keck, D. and Hiesey, W. The concept of species based on experiment. Amer. Journ. Bot. 26: 103-106, 1939.
Clausen, R. T. The terms "subspecies" and "variety". Rhodora, 43: 157-167, 1941.
Corner, E. J. H. Centrifugal stamens. Journ. Arnold Arb. 27: 423-437, 1946.
Coulter, J. M. Evolution of sex in plants. Chicago, 1914.
Lawrence 089
Croizat, L. The concept of inflorescence. Bull. Torrey Bot. Club, 70: 496-509,
1943.
_____. History and nomenclature of the higher units of classification. Bull.
Torrey Bot. Club, 72: 52-75, 1945.
De Candolle, A. P. Theorie élémentaire de la botanique. Paris, 1813.
_____. Organographie végétale. Vol. 1. Paris, 1827.
Dickson, A. Suggestions on fruit classification. Journ. Bot. 9: 309-312, 1871.
Dobzhansky, T. Genetics and the origin of species. ed. 2. New York, 1941.
Douglas, G. E. The inferior ovary. Bot. Rev. 10: 125-186, 1944.
Du Rietz, G. E. The fundamental units of biological taxonomy. Svensk Bot. Tidskr. 24: 333-428, 1930.
Eames, A. J. The role of flower anatomy in the determination of angiosperm
phylogeny. Proc. Int. Cong. Plant Sci. 1926 (Ithaca, N.Y.) 1: 423-427, 1929.
_____. The vascular anatomy of the flower with refutation of the theory of
carpel polymorphism. Amer. Journ. Bot. 18: 147-188, 1931.
Egler, F. E. The fructus and the fruit. Chron. Bot. 7: 391-395, 1943.
Erdtman, G. An introduction to pollen analysis. Waltham. Mass., 1943.
Faegri, K. Some fundamental problems of taxonomy and phylogenetics. Bot. Rev. 3: 400-423, 1937.
Florin, R. Die Coniferen des Obercarbons und des unteren Perms. Paleontographica, 85, Abt. B, pt. 2, Lief I-V, 1938-1940.
Fosberg, F. R. Subspecies and variety. Rhodora, 44: 153-157, 1942.
Goebel, K. I. E. Blütenbildung und Sprossgestaltung, Anthokladian und Infloreszenzen, 1931. [Suppl. 2 to Organographie der Pflanzen, ed. 3.]
Goethe, J. W. von. Versuch die Metamorphose der Pflanzen zu erklären. Gotha, 1790.
Goodspeed, T. H. and Bradley, M. T. Amphidiploidy. Bot. Rev. 8: 271-316, 1942.
Gray, A. Structural botany, ed. 6. New York, 1879.
_____. Gray's lessons in botany. rev. ed. New York, 1887.
Greenman, J. M. The concept of the genus. III. Genera from the standpoint of morphology. Bull. Torrey Bot. Club, 67: 371-374, 1940.
Gustafsson, Ä. Apomixis in the higher plants. Lunds Univ. Arsskrift, N.F. 42(3) and 43(2,12), 1946-1947. (For review, cf. Stebbins, G. L. in Evolution, 3: 98-101, 1949.)
Hall, H. M. The taxonomic treatment of units smaller than species. Proc. Int. Cong. Plant Sci. 1926 (Ithaca, N.Y.) 2: 1461-1468, 1929.
Hogben, L. Problems of the origins of species. In Huxley, J. The new systematics, 269-286, Oxford, 1940.
Hort, A. F. (translator). The "Critica botanica" of Linnaeus. Rev. by M. L. Green, London, 1938.
Hunt, K. W. A study of the style and stigma, with reference to the nature of the carpel. Amer. Journ. Bot. 24: 288-295, 1937.
Hutchinson, J. British flowering plants. London, 1948.
Huxley, J. S. Species formation and geographical isolation. Proc. Linn. Soc. (Lond.) 150: 253-264, 1938.
Jackson, G. The morphology of flowers of Rosa and certain closely related genera. Amer. Journ. Bot. 21: 453-466, 1934.
Johansen, D. A. Plant embryology. 305 pp. Waltham, Mass., 1950.
Jones, S. G. Introduction to floral mechanisms. 274 pp. London and Glasgow, 1939.
Just, T. The morphology of the flower. Bot. Rev. 5: 115-131, 1939.
_____. The proper designation of vascular plants. Bot. Rev. 11: 299-309, 1945.
_____. The use of embryological formulas in plant taxonomy. Bull. Torrey Bot.
Club, 73: 351-355, 1946.
[{p090}]
_____. The relative value of taxonomic characters. Amer. Midl. Nat. 36: 291-297, 1946.
Kaussmann, B. Vergleichende Untersuchungen über die blattnatur des Kelch-, Blumen-, und Staubblätter. Bot. Arch. (Leipzig), 42: 503-572, 1941.
Lamprecht, H. Systematik auf genetischer und zytologischer Grundlage. Agr. Hort. Genetica, 7: 1-26, 1949.
Link, J. H. F. Grundleheren der Anatomie und Physiologie der Pflanzen, 304 pp. Berlin, 1807, Nachträge, 84 pp., 1809.
Linnaeus, C. Fundamenta botanica ... Amsterdam, 1736.
_____. Philosophia botanica ... ; and by Hort, A., 1938. Stockholm, 1751. (Eng.
transl. by Rose, H., 472 pp. London, 1775.)
Lotsy, J. P. On the species of the taxonomist in its relation to evolution. Genetica, 13: 1-16, 1931.
MacDaniels, L. H. The morphology of the apple and other pome fruits. Cornell Univ. Agr. Exp. Sta. Mem. 230, 1940.
Maheshwari, P. The place of angiosperm embryology in research and teaching.
Journ. Indian Bot. Soc. 24: 25-41, 1945.
_____. An introduction to embryology of angiosperms. 453 pp. New York, 1950.
Masters, M. T. Classification of fruits. Nature, 5: 6, 1871.
Metcalfe, C. R. and Chalk, L. Anatomy of the dicotyledons; leaves, stem, and wood in relation to taxonomy, with notes on economic uses. 2 vols, Oxford, 1950.
Moran, R. Delimitation of genera and subfamilies in the Crassulaceae. Desert Plt. Life, 14: 125-128, 1942.
Nägeli, C. W. von. Mechanisch-physiologische Theorie der Abstammungslehre, 822 pp. München, 1884.
Ozenda, P. Researches sur les dicotylédones apocarpiques. 183 pp. Paris, 1949. [Reprinted from, Publ. Lab. de l'Ecole Normale Supérieure, Ser. Biol. vol. 2, 1949.]
Parken, J. The classification of flowering plants. Northwest Sci. 20: 18-27, 1946.
Pilger, R. K. F. Über Verzweigung und Blütenstandsbildung bei den Holzgewachsen. Bibl. Bot. 23: 1-38, 1922.
Pope, M. A. Pollen morphology as an index to plant relationship -- I. Morphology of pollen. Bot. Gaz. 80: 63-73, 1925.
Rickett, H. W. The classification of inflorescences. Bot. Rev. 10: 187-231, 1944.
Roeper, J. A. C. Observations sur la nature des fleurs et des inflorescences, Seringe Mél. Bot. 2: 71-114, 1826. [Also a Latin translation in Linnaea, 1: 433-466, 1826.]
Rutgers, F. L. Reliquae Treubianae III. Embryosac and embryo of Moringa oleifera Lam. Ann. Jard. Bot. Buitenzorg, 33: 1-66, 1923.
Sherff, E. E. The concept of the genus. IV. The delimitations of genera from the conservative point of view. Bull. Torrey Bot. Club. 67: 375-380, 1940.
Stearn, W. T. The use of the term "clone". Journ. Roy. Hort. Soc. 74: 41-47, 1947.
Stebbins, G. L. Jr. Apomixis in the angiosperms. Bot. Rev. 7: 507-542, 1941.
_____. The role of isolation in the differentiation of plant species. Biol. Symp.
6: 217-233, 1942.
_____. The genetic approach to problems of rare and epidemic species. Madroño,
6: 241-258, 1942.
Tournefort, J. P. de. Institutiones rei herbariae. Paris, 1700; ed. 3, 1719. [This work essentially a Latin edition of his Elemens de botanique. An English translation in 2 vols., probably by J. Martyn, London, 1719-1730.]
Van Tieghem, P. Recherches sur la structure du pistil et sur l'anatomie compareé de la fleur. Mém. sav. étrang. à l'inst. II, 21: 1-261, 1871. [{p091}]
Weatherby, C. A. Subspecies. Rhodora, 44: 157-167, 1942.
Wilson, C. L. The phylogeny of the stamen. Amer. Journ. Bot. 24: 686-699, 1937,
_____. The evolution of the stamen. Chron. Bot. 6: 245, 1941.
_____. The telome theory and the origin of the stamen. Amer. Journ. Bot. 29:
759-765. 1942.
Wilson, C. L. and Just, T. The morphology of the flower. Bot. Rev. 5: 97-131. 1939.
Winkler, H. Versuch eines "natürlichen" Systems der Früchte. Beitr. Biol.
Pflanzen, 26: 201-220, 1939.
_____. Zur Einigung und Weiterführung in der Frage des Fruchtsystems. Op. cit.
27: 92-130, 1940.
Wodehouse, R. P. Pollen grains, their structure, identification and
significance in science and medicine. 574 pp. New York, 1935.
_____. Evolution of pollen grains. Bot. Rev. 2: 67-84, 1936.
fn083-35. In some works (e.g., Gray's Manual, ed.7)
staminate flowers are designated neutral since they are incapable of producing
seed. However, as explained on p.84, the term neutral is reserved for flowers
lacking either sex element.
Go back fn083-35b
fn083-36. Many authors have misapplied the terms
dioecious and monoecious to flowers rather than restricting them to plants.
However, of one holds a single unisexual flower in hand, it is not possible to
determine from examination of it whether dioecism or monoecism exist; one must
know the character of all the unisexual flowers on the plant to determine which
condition exists -- hence the term applies to the plant, or to the taxon, and
not to the flower.
Go back fn083-36b
fn084-37. The terms actinomorphic and zygomorphic (or
their synonyms) are also often applied to flowers instead of only to the
perianth. Unfortunately many authors, without precise application of terms, have
described flowers as actinomorphic (or otherwise) when only the perianth (or
only the corolla) was intended.
Go back fn084-37b
fn084-38. The student should differentiate between the
terms distinct and free as used taxonomically. The term
distinct is employed to indicate complete separation of like parts (as one
petal distinct and not connate to another petal), whereas free indicates
complete freedom from union of adjoining, but unlike, parts (as the stamens free
and not adnate to the petals).
In addition to the above, and in the interest of precision of
expression, the student should understand the differences between the terms
connate, coherent, adnate, and adherent. Connate means the
fusion of like parts (as petal to petal); coherent means the meeting in
close proximity by cohesion (viscidity, vesture, etc.) or otherwise of like
parts but lacking fusion of their tissues (as pollen grains sometimes coherent);
adnate refers to the fusion of unlike parts (as stamens to petals);
adhesion refers to the coming into close contact of unlike parts but lacking
fusion of their tissues (as the indehiscent husk of Juglans adherent to
the nut).
Go back fn084-38b
fn086-39 For references to fruit classifications, cf.
entries under Dickson, Gray, Rendle, Masters, and Winkler.
Go back fn086-39b
(in alphabetical order)
also diecious adj. Botany 1. Having the male and female reproductive organs borne on separate individuals of the same species. 2. Characterized by species in which the male and female reproductive organs occur on different individuals; sexually distinct. [From New Latin Dioecia former class name di- 1 Greek oikia a dwelling; See weik- 1 in Indo-European Roots.] -- AHTDdioecious
Go back dioecious-b
Dry fruits: Achene; Follicle; Legume; Loment; Nut; Pyxis; Samara; Schizocarp; Utricle;

Go back Dry-fruits-b
Fleshy fruits: Berry, Drupe, Hesperidium, Pepo, Pome

Go back Fleshy-fruits-b
monoecious also monecious adj. 1. Botany Having unisexual reproductive organs or flowers, with the organs or flowers of both sexes borne on a single plant, as in corn and pines. 2. Zoology Hermaphroditic. [New Latin Monoecia class name mono- Greek oikia dwelling; See weik- 1 in Indo-European Roots.] -- AHTD
Go back monoecious-b
End of TIL file