Update:
2012-12-31 06:22 PM +0630
TIL
TAXONOMY OF VASCULAR PLANTS: Part One
04.2 Criteria of classification
criteria.htm
by George H. M. Lawrence, Professor of Botany
at the Bailey Hortorium, Cornell University, 1951
Photo-copied by Maung Kan Tun from the original text.
Edited and set in HTML by U Kyaw Tun (M.S., I.P.S.T., USA),
Tun Institute of Learning (TIL) and the staff of TIL.
Not for sale. No copyright. Free for
everyone. Prepared for students and
staff of TIL Computing and
Language Center, Yangon, MYANMAR :
http://www.tuninst.net ,
http://www.softguide.net.mm ,
www.romabama.blogspot.com
index.htm |
Top
Lawrence-indx.htm
Principles of Taxonomy
Criteria of classification
- Morphological criteria p056
Vegetative vs
reproductive structures p057
Ligneous vs.
herbaceous characters p058
Inflorescences p059
Evolution of inflorescences
p060
Fig. 3. Schematic diagrams of hypothetical evolution
of inflorescence types 063
Foot notes
UKT notes
Vegetative structures
Contents of this page
[{cont. p056}]
All taxonomists are agreed that the differences between plants, and the
similarities that plants may possess in common, are measurable to a large degree
by the morphological characters of those plants. A morphological character, to a
taxonomist, is one inherent in or manifested by
Lawrence 057
a structural component of the plant. In general it is concerned with the
organography of the plant, with characters that usually are discernible with the
aid of no more than a good hand lens. The more recent recognitions by
taxonomists of the significance of the related sciences of anatomy, cytology,
and comparative morphology have resulted in an expansion of the concept as to
what is a morphological character and to the extreme that some taxonomists now
consider the number of chromosomes to represent as significant a morphological
character as that provided by the number of
stamens. The value of the
morphological character is measured by its constancy. The more constant the
character, the greater is the reliability that can be placed upon it. In this
regard it must be remembered that a character that is reasonably constant among
the members of one group may be a weak character among members of another group.
For example, among the families of some dicotyledonous plants the character of
ovary position (inferior vs. superior) is adequate to separate families and
orders, but within some monocotyledonous plants the two types of ovary position
are to be encountered within a family (e.g., Bromeliaceae), or even within a
single genus (e.g., Bomarœa).
Contents of this page
Vegetative vs. reproductive
structures.
The vegetative structures of plants (i.e., those of leaves,
stems, buds, and habit of growth) are readily apparent to the eye and have an
appeal to the beginning taxonomist. Their observation generally does not require
the use of a microscope or high-powered lens nor the employment of razor blade
or dissecting needles. However, their use as taxonomic criteria is limited for
two basic reason: the total number of vegetative characters available is few as
compared with the number of species of vascular plants, and too often the
vegetative characters are not particularly constant.
The number of species of woody plants of any temperate flora is usually fewer
than the number of species of herbaceous plants of the same flora. For this
reason, most genera, and sometimes the species, of these woody plants may be
separated largely and almost exclusively by vegetative characters. Keys to
woody plants are popular for this reason. However, even among the genera of
woody plants, vegetative characters lack the necessary variability of type,
stability, and constancy necessary for the identification of species. Examples
are abundant in such temperate genera as Crataegus, Prunus,
Quercus, Rhododendron, Salix, and Vaccinium. As a rule,
the greater the number of species in any genus of woody plants the more
difficult is their identification by means of vegetative characters.
Lawrence 058
In the preparation of keys13
(fn058-13)
to woody plants it frequently is desirable to use vegetative characters to as
great an extent as practicable. It has been found from experience, in dealing
with plants of temperate regions, that certain vegetative characters are
generally more constant than are others. (In certain genera and families,
however, there are notable exceptions.) Those characters known to be more
reliable than others are listed below in sequence of approximate descending
constancy:
Leaves needle- or scalelike vs. leaves broad and laminate
Venation parallel vs. venation reticulate (netted)
Leaves compound vs. leaves simple
Plants erect trees or shrubs vs. plants vinelike
Stipules present vs. stipules absent
Leaves lobed or divided vs. leaves neither lobes nor divided
Leaves opposite or whorled vs. leaves alternate
Leaf margins entire vs. leaf margins variously toothed
Leaves persistent and
coriaceous vs. leaves deciduous
Spines or prickles present vs. spines or prickles absent
Leaf base
tapering (cuneate) vs. leaf base broad (cordate, obtuse, truncate)
(UKT: see leaf shapes )
Leaves with 1 main mid-vein vs. leaves with 3 primary veins
Succeeding the above would be characters of leaf form, leaf apex, the leaves
petioled or sessile, and pith characters of the twig or stem. Characters of
vesture (pubescence), surface, coloration, milky vs. watery sap, and sizes of
plant or leaves are generally useful only in the separation of species or their
variants.
See UKT note on vegetative
structures adapted from on Trees Commonly Cultivated in Southeast Asia, an illustrated
field-guide, 2nd ed., FAO, 1999, pp.230
Reproductive structures are those associated with the flower and fruit. The
number of organs in these structures, and the number of components of these
organs, are many times that available from vegetative structures. For this
reason the number of mathematical combinations and recombinations of
reproductive characters is infinitely greater and ordinarily exceeds the number
of species (or smaller units) to be differentiated. In addition to being more
abundant, reproductive characters in general are much more nearly constant than
are vegetative characters. The reliability of any single reproductive character
selected for taxonomic use must be determined individually, however, for each
particular group of plants.
Contents of this page
Ligneous vs. herbaceous
characters
In recent decades increased emphasis has been placed on
the phyletic significance of the woody as opposed to the herbaceous character of
plants. It is believed by many botanists that among the higher vascular plants,
those of woody habit were the antecedents of those of herbaccous habit.
Hutchinson, in the latest version of his classification (1948), has made this
hypothesis the
Lawrence 059
basis of his two classes (Lignosae and Herbaceae) of the
Dicotyledoneae. In doing so he warned
. . . against taking the view that he has reverted to the old practice of
putting all the woody plants into one group and all the herbaceous plants into
another. . . . what has been done is to put all those families which are
predominately woody, and the more primitive genera of which are
woody, into the woody phylum, and conversely the same has been done with the
herbaceous families.
In other words, the woody division consists of trees and shrubs plus
herbs which are related to trees and shrubs, and the herbaceous
division consists of herbs plus some clearly related woody plants.
In addition to the significance placed on these criteria by Hutchinson, the more
elemental distinction between woody and herbaceous plants has practical value in
artificially separating the minor units of classification. In many cases, the
demarcation between the two criteria is not sharp. Some plants that appear to be
herbaceuos are actually woody in that their stems in addition to being woody in
texture are of perennial duration (e.g., Gaultheria procumbens, Epigaea repens,
or Vinca minor). Other plants are basally woody but have stems that for the most
part are herbaceous. Such plants are often referred to as being suffrutescent in
character (e.g., Pachysandra terminalis, Arabis caucasica, or
Aralia hispida).
Contents of this page
Inflorescences
An inflorescence
conventionally is considered to be the arrangement of flowers on the floral
axis. By Linnaeus (1751), it was said to be a mode of flowering; that is, he
considered it to be a phenomenon or function and not a structural component of
the plant. The present-day classification of inflorescences, although utilizing
to an extensive degree the terminology of Linnaeus, is based on the relatively
unscientific observations of Link (1807) as modified by Roeper (1826)14
(fn059-14) These botanists
considered the inflorescence to be a structural entity of the plant, subject to
subdivision into definite classes and types for use as taxonomic criteria. The
basic classification of the inflorescence by Roeper (into a series of
determinate and indeterminate types) is, with some amplifications and
modifications, that used today in taxonomic and less technical descriptions of
flowering plants.15 (fn059-15)
As pointed out by Rickett (p.189), inflorescences are branch systems and are "all related
Lawrence 060
in the sense of being derivable from one underlying type." It is conceded
generally that the primitive inflorescence or its antecedent was a system of
branches each of which was terminated by a flower. Every individual flower is a
terminal unit. It is borne on the end of an axis, and it matters not whether
that axis conventionally be termed a peduncle, pedicel, receptacle, torus,
or stem. The structure represented by any one of these terms is technically a
floral axis. It is important to an understanding of the use of inflorescences in
the classification of these plants to know what type of inflorescence is
primitive, and of the possible developmental sequence of other types from the
primitive. Unfortunately there is yet insufficient knowledge concerning the
phyletic relationships of the families of flowering plants to permit more
than speculation on the origins of inflorescences. However, speculations based
on what are believed to be the best available data do provide rational
approaches to a solution of the problem.
Contents of this page
Evolution of inflorescences
 Three theories have been proposed to account for the evolution of present-day
inflorescences from the presumed primitive condition of a branch system of
terminal flowers. The first postulates the primitive type to have been a
panicle, the second that it was the single terminal flower,
Three theories have been proposed to account for the evolution of present-day
inflorescences from the presumed primitive condition of a branch system of
terminal flowers. The first postulates the primitive type to have been a
panicle, the second that it was the single terminal flower,
 and the third that
it was a dichasium. The first theory has been held by many investigators,
including Nägeli (1883), Celakovský (1892), and Pilger (1922). By acceptance of
the panicle as primitive, it is easy to explain the derivation of all
other types from it as was done in part by Goebel (1931). Pilger supported
his views in defense of the theory with analogies which he conceded do not exist
among living woody angiosperms. The second theory, supported by Parkin (1914)
proposes the development of multiflowered inflorescences from the solitary
flower by addition of lateral flowers (and branches of flowers) by axillary
growth. Any consideration of this theory must take into account the generally
accepted view, discussed later in his chapter, that branch systems are a
fundamental part of the plant organization, and that the flower is probably made
up of highly modified branch systems. Acceptance of the belief that foliar
organs are highly modified branch systems adds logic to a theory that the primitive
inflorescence also was a branch system and that most existing examples of
solitary flowers
and the third that
it was a dichasium. The first theory has been held by many investigators,
including Nägeli (1883), Celakovský (1892), and Pilger (1922). By acceptance of
the panicle as primitive, it is easy to explain the derivation of all
other types from it as was done in part by Goebel (1931). Pilger supported
his views in defense of the theory with analogies which he conceded do not exist
among living woody angiosperms. The second theory, supported by Parkin (1914)
proposes the development of multiflowered inflorescences from the solitary
flower by addition of lateral flowers (and branches of flowers) by axillary
growth. Any consideration of this theory must take into account the generally
accepted view, discussed later in his chapter, that branch systems are a
fundamental part of the plant organization, and that the flower is probably made
up of highly modified branch systems. Acceptance of the belief that foliar
organs are highly modified branch systems adds logic to a theory that the primitive
inflorescence also was a branch system and that most existing examples of
solitary flowers
 represent examples of reduction and suppression. The third
theory, that the dichasium (Fig. 3)*
may represent the primitive type, has been accepted by Rickett (1944).
represent examples of reduction and suppression. The third
theory, that the dichasium (Fig. 3)*
may represent the primitive type, has been accepted by Rickett (1944).
*UKT: For reader's convenience this place is bookmarked as fig3call1b.
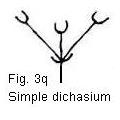
In its simplest form the primitive dichasium consisted of three flowers disposed
in a cluster formed on a single peduncle by a dichotomous branching immediately
beneath a terminal flower (Fig. 3a). From this
Lawrence 061
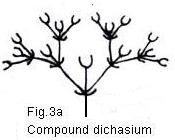
simple dichasium there may be developed a more complex dichasium that is
formed by "a repetition of the same apparent dichotomy in each lateral branch."
In nature, the dichasium may be simple or compound, ample or restricted, and it
would be hazardous to speculate as to which form preceded the other in ancestral
types. There is no clear paleobotanical evidence to support the view that the
dichasium was ancestral to the panicle, neither can the latter be established as
the more primitive. Either one may have been derived from the other. There is
some fossil evidence that in a few rare instances the solitary flower may
represent a primitive situation. It has been shown (Florin, 1948) that
primitive and now extinct conifers possessed a paniculate inflorescence, a fact
that should not, however, be construed as evidence indicative of gymnospermous ancestry of
the angiosperms. There is a theory held by many botanists that the
Pteridospermae (seed plants with fernlike foliage) provide the ancestral group
best possessing angiospermlike morphology; in this taxon the ovules, borne on
complex fernlike leaves, suggest a paniculate arrangement. Irrespective of the
relative primitiveness of the panicle and the dichasium, the views of Rickett on
the possible evolution of inflorescence types, as outlined below, are most
helpful and provide a logical understanding of possible relationships.

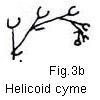
 The dichasium was postulated by Rickett to have given rise, by only
slight structural modifications, to inflorescence types such as bostryches, cincinni,
verticels, cymes, helicoid cymes, scorpioid cymes, and by apparent elimination
of internodes it may have
The dichasium was postulated by Rickett to have given rise, by only
slight structural modifications, to inflorescence types such as bostryches, cincinni,
verticels, cymes, helicoid cymes, scorpioid cymes, and by apparent elimination
of internodes it may have
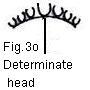 been an antecedent to types such as the umbel,
capitulum [UKT: head], and corymb
(Fig. 3).
The obvious structural organization of
complex and congested inflorescences indicates
been an antecedent to types such as the umbel,
capitulum [UKT: head], and corymb
(Fig. 3).
The obvious structural organization of
complex and congested inflorescences indicates
 that many of them may have developed
from various combinations and aggregations of dichasia. In this regard, it is
now believed that the seemingly simple raceme and spike
represent types that are reduced, along one of two or more evolutionary lines, from
more complex ancestral types of dichasia or panicles. That is, the raceme or the spike may
have been evolved through steps involving the reduction of individual dichasia
to as little as a single flower
that many of them may have developed
from various combinations and aggregations of dichasia. In this regard, it is
now believed that the seemingly simple raceme and spike
represent types that are reduced, along one of two or more evolutionary lines, from
more complex ancestral types of dichasia or panicles. That is, the raceme or the spike may
have been evolved through steps involving the reduction of individual dichasia
to as little as a single flower

 accompanied or not by similar reduction of leaves to subtending bracts. It has been established that, in many cases, the
presumed distinction of an inflorescence being determinate (i.e., centripetal)
or indeterminate (centrifugal) is without validity and that ultimately in
a new classification of inflorescence types, this criterion will have less value
than is currently accorded it. In certain inflorescence types it is a valid
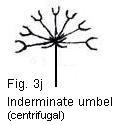
characteristic, while in others it is unreliable. For example, the umbel of
members of the Umbelliferae is an indeterminate
accompanied or not by similar reduction of leaves to subtending bracts. It has been established that, in many cases, the
presumed distinction of an inflorescence being determinate (i.e., centripetal)
or indeterminate (centrifugal) is without validity and that ultimately in
a new classification of inflorescence types, this criterion will have less value
than is currently accorded it. In certain inflorescence types it is a valid
characteristic, while in others it is unreliable. For example, the umbel of
members of the Umbelliferae is an indeterminate
Lawrence 062
type of inflorescence whose outer flowers open before the inner ones. However,
in many of the Amaryllidaceae (as in
Allium 16
(fn062-16)) the
umbel is a determinate inflorescence as indicated by the central flower opening prior to the
outer flowers. Structurally, the inflorescence is an umbel in either family, but phyletically
the two kinds of umbel are unrelated and represent reductions by two different
evolutionary lines (Fig. 3).
A derivative and advanced group of inflorescence
types over those considered to have more direct affinity with the dichasium is that
known as the monochasium. A monochasium is an inflorescence resulting from a dichasium
in which one branch of each dichotomy continues to develop while the other
branch is suppressed completely (e.g., aborts). The result is a
sympodial axis
composed of series of superposed axis and is sometimes referred to as a sympodial dichotomy
(Fig.3). This
 type of inflorescence is phyletically a
complex inflorescence, since it is made up of a series of branch systems. It is
the "scorpioid cyme" of botanical floras and manuals and is characteristic of
many boraginaceous genera (Myosotis, Heliotropium, Symphytum)
as well as of representatives of the
type of inflorescence is phyletically a
complex inflorescence, since it is made up of a series of branch systems. It is
the "scorpioid cyme" of botanical floras and manuals and is characteristic of
many boraginaceous genera (Myosotis, Heliotropium, Symphytum)
as well as of representatives of the
 Polemoniaceae and Hydrophyllaceae.17
(fn062-17)
[
Polemoniaceae and Hydrophyllaceae.17
(fn062-17)
[ {hsing-nha-maung:kri:}
Heliotropium indicum L. , Family: Boraginaceae.]
In these taxa its character and origin are usually obvious by macroscopic inspection.
However, reduction often has advanced to a degree where the monochasium is not as
apparent in all cases as in the so-called scorpioid cyme. That is, in some groups,
the sympodial false
{hsing-nha-maung:kri:}
Heliotropium indicum L. , Family: Boraginaceae.]
In these taxa its character and origin are usually obvious by macroscopic inspection.
However, reduction often has advanced to a degree where the monochasium is not as
apparent in all cases as in the so-called scorpioid cyme. That is, in some groups,
the sympodial false
 axis(actually composed of determinate units) may be so refined in
appearance as to seem to be a simple and true axis of indeterminate growth; such an
inflorescence is the raceme of Claytonia, an
axis(actually composed of determinate units) may be so refined in
appearance as to seem to be a simple and true axis of indeterminate growth; such an
inflorescence is the raceme of Claytonia, an
 infloresence that morphologically is as much a helicoid cyme as is that of Myosotis.
In other genera, the monochasium has been reduced to an umbel as in the florist's geranum
(Pelargonium). The umbell of Allium, mentioned above, is of similar origin.
From this brief review of possible origins of inflorescences, it may be speculated that either
the panicle or the dichasium (rarely the solitary flower) is the primitive type of
inflorescence, and that from them have been evolved all other types of
inflorescences. That is, the solitary flower, the panicle, and the dichasium all
may be primitive types.18(fn062-18)
By this speculation,
infloresence that morphologically is as much a helicoid cyme as is that of Myosotis.
In other genera, the monochasium has been reduced to an umbel as in the florist's geranum
(Pelargonium). The umbell of Allium, mentioned above, is of similar origin.
From this brief review of possible origins of inflorescences, it may be speculated that either
the panicle or the dichasium (rarely the solitary flower) is the primitive type of
inflorescence, and that from them have been evolved all other types of
inflorescences. That is, the solitary flower, the panicle, and the dichasium all
may be primitive types.18(fn062-18)
By this speculation,
Lawrence 063
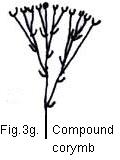
Fig. 3. Schematic diagrams of hypothetical evolution of inflorescence types

Fig. 3. Schematic diagrams of hypothetical evolution of inflorescence types:
a. compound diachasium; b. helicoid cyme; c. cincinnus;
d. scorpioid cyme; e. thyrse; f. panicle;
g. compound corymb; h. simple corymb; i. raceme;
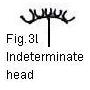
j. indeterminate umbel; k. spike; l. indeterminated head;
m. cyme; n. determinate umbel; o. determinate head;
p. verticillate inflorescence; q. simple dichasium.
Go back to fig3call1b/
fig3call2b/ fig3call3b/
fig3call4b
Lawrence 064
it is indicative that usually the conventional raceme is more
primitive than the spike but that both are advanced over the panicle, that while
 the panicle may have been primitive, some present-day examples of the panicle
may have been derived from a thyrse, and that the numerous cymose types of
inflorescences are perhaps more primitive than most of those mentioned above.
Accordingly, the modern or present-day inflorescence represented by a solitary
flower in almost all plants is a product of reduction, a reduction that may have
resulted from any one of many multiflowered inflorescence type, whose origin may be
indicated only by comparative studies of infloresences of related taxa.
the panicle may have been primitive, some present-day examples of the panicle
may have been derived from a thyrse, and that the numerous cymose types of
inflorescences are perhaps more primitive than most of those mentioned above.
Accordingly, the modern or present-day inflorescence represented by a solitary
flower in almost all plants is a product of reduction, a reduction that may have
resulted from any one of many multiflowered inflorescence type, whose origin may be
indicated only by comparative studies of infloresences of related taxa.
The significance of the inflorescence as a taxonomic criterion has been
established. In some families it is of significance in recognizing entire
families (Umbelliferae, Cornaceae, Compositae, Labiatae, Gramineae,
Amaryllidaceae [sensu Hutchinson]) and in others in recognizing tribes or genera
(Galium, Smilax, Malus, Philadelphus). The consideration of phyletic
significance of the inflorescence cannot be evaluated apart from other criteria,
but in dealing with minor units of flowering plants often it may be helpful in
establishing their position in a particular schema or classification.
Contents of this page
fn058-13. Cf. pp. 225- 228 for discussion of keys,
their preparation, and use.
Go back fn058-13b
fn059-14. See Rickett, H. W. (1944), for a detailed
analysis of the subject of inflorescence classification, and from whose review
much of the data for this section of the chapter were obtained. See also Croizat
(1944) for views treating the inflorescence as an aggregate of vegetative and
floriferous axes.
Go back fn059-14b
fn059-15. A determinate inflorescence, simply
expressed, is one in which the terminal flower of the axis opens first with the
remaining flowers of that axis opening successively from top to bottom. An
indeterminate inflorescence is one in which the lowermost flowers of an axis
open first and the terminal flowers open last, with the successive opening of
flowers accompanied by an elongation of the axis.
Go back fn059-15b
fn062-16. For discussion of the family to which
Allium belongs (Liliaceae vs. Amaryllidaceae), cf. Part II, p. 415.
Go back fn062-16b
fn062-17. Note from study of Fig. 3, That the so-called
scorpioid cyme here referred to in quoted is properly a helicoid cyme (helicos,
mening , snaillike) and that the true scorpioid cyme is an inflorescence type
not commonly encountered.
Go back fn062-17b
fn062-18. The pteridosperm leaves beat ovules in ways
that would provide a background and historical basis for this: the ovule
terminal and solitary on huge fronds; the ovules terminal on few or all pinnules;
or the ovules marginal and terminal throughout the
Go back fn062-18b
Contents of this page
stamen (def.)
stamen n. pl. stamens or
stamina 1. The pollen-producing reproductive organ of a flower, usually
consisting of a filament and an anther. [Latin stāmen thread] -- AHTD
Go back stamen-def-b
capitulum
capitulum n. pl. capitula 1.
Botany See flower head. -- AHTD
head n. 20. Botany A flower head. -- AHTD
flower head n. 1. Botany A
dense, short, compact cluster of sessile flowers, as of composite plants or
clover. Also Called capitulum . 2. A very dense grouping of flower buds,
as in broccoli and cauliflower. -- AHTD
Go back capitulum-b
Contents of this page
sympodial axis
sympodium n. pl. sympodia Botany 1.
A primary axis that develops from a series of short lateral branches and often
has a zigzag or irregular form, as in orchids of the genus Cattleya.
Also Called pseudaxis . -- AHTD
Go back sympod-axis-b
Contents of this page
stipule
stipule n. Botany 1. One of the usually small,
paired appendages at the base of a leafstalk in certain plants, such as roses
and beans. [New Latin stipula from Latin stalk] -- AHTD
Go back stipule-b
Contents of this page
coriaceous
coriaceous adj. 1. Of or like leather, especially
in texture. [From Late Latin cori³ceus from Latin corium
leather] -- AHTD
Go back coriaceous-b
Contents of this page
Vegetative structures
For identification of a plant or differentiation between two similar plants
in field-work, vegetative structure are more important than reproductive
structures: the primary reason being vegetative structures can be observed all
year round whereas reproductive structures can be observed only for a short
period. Among the vegetative structures, the character of the leaves seems to be
the most important :
 1.
Arrangement of leaves:
1.
Arrangement of leaves:
opposite, alternate, whorled
2. Leaf structures:
simple, palmately compound, even pinnately
 compound, odd-pinnately compound, bipinnately compound.
(pinnate also pinnated adj. 1.
Resembling a feather; having parts or branches arranged on each side of a common
axis: a polyp with a pinnate form; pinnate leaves. [Latin pinn³tus
feathered from pinna feather; See pet- in Indo-European Roots.] -- AHTD)
compound, odd-pinnately compound, bipinnately compound.
(pinnate also pinnated adj. 1.
Resembling a feather; having parts or branches arranged on each side of a common
axis: a polyp with a pinnate form; pinnate leaves. [Latin pinn³tus
feathered from pinna feather; See pet- in Indo-European Roots.] -- AHTD)
 3.
Leaf shapes:
3.
Leaf shapes:
needle, scalelike, linear, lanceolate, oblong, elliptical, spatulate,
ovate, orbicular, reniform, cordate, obovate, cuneate, oblanceolate, falcate
(Go back leaf-shape-b)
4. Leaf margins:

entire, sinuate, serrate, dentate, lobed, clefted, crenate, incised, parted, divided
 5.
Leaf tips:
5.
Leaf tips:
acuminate, acute, obtuse, rounded, truncate, emarginate
6. Leaf bases:
cuneate, acute, obtuse, rounded, truncate, auricula (auricle n.
2.
 Biology An earlobe-shaped part, process, or appendage, especially at the base of
an organ. -- AHTD).(Go back leaf-base-b)
Biology An earlobe-shaped part, process, or appendage, especially at the base of
an organ. -- AHTD).(Go back leaf-base-b)
 7. Leaf venation:
7. Leaf venation:
pinnate, arcuate, parallel, palmate
Go back to
veg-structures-b
Contents of this page
determinate
determinate adj.
4. Botany a. Terminating in a flower and blooming in a
sequence beginning with the uppermost or central flower; cymose: a
determinate inflorescence. b. Not continuing indefinitely at the tip
of an axis: determinate growth. -- AHTD
Go back determin-b
Contents of this page
End of TIL file